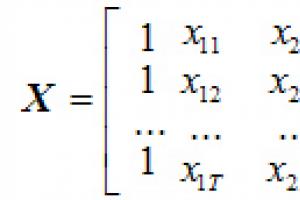Oblici citokroma p450. P450 citokroma. Geni ljudskog citokroma P450
Polunina T.E.
Oksana Mihajlovna Drapkina
– Nastavljamo s našim programom. Naša predavanja i rasprave o ginekologiji su pri kraju, potpuno smo ušli u propise, pa ćemo se truditi da ih ne napustimo. Profesorica Tatyana Evgenievna Polunina otvara odjel za gastroenterologiju. Predavanja “Uloga obitelji citokroma P450 u patogenezi i liječenju nealkoholne masne bolesti jetre.”
Tatyana Evgenievna Polunina, prof., doktor medicinskih znanosti:
– Citokromi P450 (CYP 450) naziv je velike obitelji univerzalnih enzima u ljudskom tijelu. Citokromi P450 igraju važnu ulogu u oksidaciji brojnih spojeva, kao što su endogeni spojevi (steroidi, žučne kiseline, masne kiseline, prostaglandini, leukotrieni, biogeni amini), kao i egzogeni spojevi (lijekovi, industrijsko onečišćenje, pesticidi, kancerogeni i mutageni) ), potonji se nazivaju ksenobiotici.
Na ovom slajdu možete vidjeti gdje se nalaze citokromi P450. Nalaze se u hepatocitu, u citosolu. Osnova za lokaciju je endoplazmatski retikulum. Konkretno, lipidna membrana, koja sadrži dvosloj fosfolipida, na sebi ima nekoliko povezanih struktura. Ovo je citokrom, koji uključuje željezni protein, nikotinamid adenin dinukleotid i oksidoreduktazu, koja je uključena u metabolički kompleks lijekovi i gore prikazani ksenobiotici.
Najčešći predstavnici ove skupine kojima se kliničari obraćaju su citokromi P452 AC, P450 2D, P450 2E1, P450 3A4. Ovi enzimi kataliziraju širok raspon metaboličkih reakcija i jedan citokrom može metabolizirati nekoliko lijekova koji imaju različite kemijske strukture. Isti lijek ima različite učinke na citokrom P450 i na različite organe. A posebno najvažniji citokrom na koji obraćamo pažnju je citokrom P450 2E – najvažniji izoenzim citokroma P450, razgrađuje lipoproteine niske gustoće.
Trenutno nisu razvijene samo metode fenotipizacije koje se temelje na specifičnosti supstrata određenih izoenzima citokroma P450, već i aktivnost određenog enzima i metabolizma određuju farmakokinetika supstrata markera i promjene u koncentracijama nepromijenjene tvari. i njegov metabolit. Ali određivanje izoenzima citokroma P450 identificiranjem gena za odgovarajuće izoenzime provodi se pomoću lančane reakcije polimeraze. To se naziva genotipizacija izoenzima citokroma P450.
Na ovom slajdu vidimo da se u hepatocitu nalazi mjesto gdje se nalazi endoplazmatski retikulum, citokromi P450, kojih ima više od 50, te lijekovi koji se razgrađuju u određenom citokromu, u nekim slučajevima spaja se s citokromom i formira vezikula koja oštećuje hepatocit, uzrokujući istovremeno stres i citokine; dovodi do aktivacije čimbenika nekroze tumora, a posebno je okidač za pokretanje kaspaza, što se manifestira katalitičkim procesima.
Nealkoholna masna bolest jetre, koja je naknadno identificirana kao nozološki entitet, počela se nazivati nealkoholna masna bolest jetre (NAFLD) od 1980. godine, nakon otkrivanja promjena na jetri pacijenata nealkoholičara koje su bile slične onima viđenim u oštećenja izazvana alkoholom.
Prirodna povijest nealkoholne masne bolesti jetre uključuje steatozu kao početnu fazu, koja, bez napredovanja, može biti asimptomatska, te steatohepatitis, koji je praćen strašnim vegetativnim manifestacijama, sindromom citolize i dispeptičkim manifestacijama. S razvojem fibroze javlja se prilično ozbiljan problem - ciroza jetre, a zatim se razvija portalna hipertenzija i karcinom.
Želio bih vam skrenuti pozornost na činjenicu da je još 1894. Kiernan predložio određenu arhitekturu jetre, koja se sastoji od gredne strukture. Na periferiji greda, koje se sastoje od poligonalnih hepatocita, nalazi se trijada: žučni kanal, portalna vena i arterija. Ovaj slajd predstavlja normalnu zdravu jetru i masnu infiltraciju hepatocita. U ovom dijagramu morfološki je prikazana steatoza jetre, koja je jedna od prvih faza razvoja nealkoholne masne bolesti jetre.
Sljedeća opcija za razvoj upalnog procesa, koji dovodi do širenja fibroznog tkiva po cijeloj jetri, vidimo steatohepatitis i naknadno cirozu jetre s razvojem portalne hipertenzije. Najčešće se radi o mikronodularnoj cirozi jetre koja se već dosta jasno utvrđuje u fazama razvoja nealkoholne masne bolesti jetre, prati je portalna hipertenzija, proširene vene jednjaka, želuca, komplikacije tipične za ciroza jetre i smrt.
Uz nealkoholni steatohepatitis, najčešći razvoji su oni koji su najčešće povezani kao popratne bolesti: dijabetes, pretilost. U bolesnika se nealkoholni steatohepatitis razvija do 75%, a ako se kombinira dijabetes melitus i pretilost, tada 90% bolesnika ima nealkoholnu masnu bolest jetre.
Jetra je nedvojbeno glavni ciljni organ zahvaćen metaboličkim sindromom. Inzulinska rezistencija ključna je značajka koja je osnova za intrahepatocitno nakupljanje lipida, masnu jetru, nealkoholni steatohepatitis i cirozu jetre.
Želio bih skrenuti pozornost da metabolički sindrom ne uključuje samo poremećenu toleranciju glukoze, već i dislipidemiju, abdominalno-visceralnu pretilost, inzulinsku rezistenciju i hiperinzulinemiju, arterijsku hipertenziju, ranu aterosklerozu, poremećenu hemostazu, hiperurikemiju, hiperandrogenizam. Htio bih reći da je nealkoholna masna bolest jetre, steatoza, dio metaboličkog sindroma i trenutno je kvintet koji se nekada nazivao “smrtonosnim kvartetom”.
Čimbenici rizika prikazani na ovom slajdu ponekad se mijenjaju ovisno o različite zemlje Konkretno, američki stavovi i europski stavovi malo se razlikuju. No, ipak, opseg struka, razine triglicerida, lipoproteina, krvni tlak, posebno 130/85, razina glukoze su pokazatelji koje je potrebno pratiti kod bolesnika s metaboličkim sindromom.
Bolesti povezane s metabolizmom lipida su: nealkoholna masna bolest jetre, dijabetes melitus tipa 2, koronarna bolest jetre, hipertenzija.
U shemi patogeneze osobitu važnost ima inzulinska rezistencija masnog tkiva. Povećanje lipogeneze, odnosno povećanje razine masnih kiselina, povećanje sinteze triglicerida i lipotoksičnost dovode do razvoja inzulinske rezistencije, a to dovodi do metaboličke disfunkcije, stresa endoplazmatskog retikuluma, u kojem se također dolazi do metabolizma masnih kiselina, a posebno lipoproteina, te do aktivacije upale . To su Kupfferove stanice i zvjezdaste stanice, koje dalje dovode ne samo do povećanja razine lipida vrlo niske gustoće, nego nedvojbeno to dovodi do razvoja steatohepatitisa s fibrozom, te dolazi do aktivnosti procesa koji ide prema cirozi. od jetre.
Na razini hepatocita, masne kiseline prolaze kroz esterificiranje u trigliceride i izvoze se kao lipoproteini niske gustoće, što je situacija u normalnim hepatocitima koja je povezana s oksidacijom u mitohondrijima, peroksisomima i mikrosomima.
Bez sumnje, u mehanizmu inzulinske rezistencije, koji je ovdje prikazan, ključnu ulogu imaju faktor nekroze tumora, slobodni radikali, leptin, masne kiseline i pojačana lipoliza, što dovodi do apsorpcije masnih kiselina, do poremećaja β- oksidacije masnih kiselina u mitohondrijima i također do nakupljanja masnih kiselina u hepatocitu.
Indukcija citokroma P450 4A11 i P450 2E1 dovodi do peroksidacije lipida, što nedvojbeno dovodi do aktivacije čimbenika povezanih s nakupljanjem triglicerida. Hiperinzulinemija je ključni čimbenik koji dovodi do inzulinske rezistencije. Također dovodi do povećanja glikolize, sinteze masnih kiselina i nakupljanja triglicerida u hepatocitima.
Sljedeći slajd prikazuje mehanizam interakcije između mikrosomske oksidacije i mitohondrijske β-oksidacije. Imajte na umu da mitohondrijska Ω-oksidacija i mitohondrijska β-oksidacija dovode do aktiviranja takozvanih peroksisomalnih β-oksidacijskih receptora, a posebno receptora aktiviranih proliferatorom peroksisoma. To dovodi do izražaja akumulacije određenog proteina i sukladno tome acetil-koenzima A, koji se akumulira i pokreće mehanizam koji dovodi do preopterećenja dikarboksilnih masnih kiselina.
Na sljedećem slajdu vidite da se steatohepatitis i fibroza formiraju u pozadini mitohondrijskih reaktivnih vrsta kisika. Ključna točka Za poticanje fibroze nedvojbeno je nakupljanje malondialdehida, što dovodi do stvaranja upalnih infiltrata, fibroze i aktivacije zvjezdastih stanica. Zvjezdaste stanice pokreću indukciju citokina kao što su faktor tumorske nekroze i transformirajući faktori rasta. Iscrpljenost antioksidativnog sustava dovodi do pokretanja Fas-leganda, mitohondrijske reaktivne vrste kisika, dolazi do nekroze hepatocita, a zatim se razvija fibrozno tkivo koje je osnova za razvoj ciroze.
Ovaj slajd prikazuje dijagram; vidite višak lipida koji se nakupljaju u hepatocitu. Mitohondrijska disfunkcija i disfunkcija citokroma P450 dovodi do aktivacije peroksidacije lipida, pokretanja Kupfferovih stanica, upalnih citokina, aktivacije zvjezdastih stanica i apoptoze, što posljedično dovodi do razvoja nekroze hepatocita.
Metabolički sindrom je vrlo važan jer je nealkoholna masna jetra dio metaboličkog sindroma. I to ne samo na hepatocitu, u kojem dolazi do porasta razine lipoproteina niske i vrlo niske gustoće, triglicerida (ovo je vrlo važno), nego i na endotelnoj stanici. Dolazi do disfunkcije endotela i također se pokreće trenutak koji je povezan s peroksidacijom lipida, nakupljanjem tvari koje utječu na aterosklerozu, iznenadnu smrt i srčani udar.
Nedvojbeno je povećanje razine slobodnih masnih kiselina povezano s adipocitima. A posebno smanjenje esterificiranog kolesterola također dovodi do različitih stresova nuklearnog receptora. A takozvani aktivirani peroksisomski proliferatorski receptor trenutno je posebno važan, na njega je usmjerena sva pozornost znanstvenika koji se bave pretilošću, dijabetesom i nealkoholnom masnom bolešću jetre.
Monocit (makrofag), u nekim slučajevima, povećanjem razine upalnih odgovora (tumorski nekrotični faktor, interleukini-6, membranski toll-like receptori, slobodne masne kiseline) također pokreće događaje koji su posebno povezani s patološkim učincima masnih kiselina. kiseline.
Kriteriji za procjenu inzulinske rezistencije svima su poznati od 1985. godine. Određuje se HOMA indeksom - Homeostasis Model Assessment, te modernijim QUICKI indeksom - Quantitave Insulin Sensitivity. Ovdje su prikazane koncentracija inzulina, glukoza u serumu i norme.
Napominjemo da ne trebaju svi bolesnici s nealkoholnom masnom bolešću jetre biopsiju jetre. Trenutno imamo točke koje nam omogućuju određivanje razine masne infiltracije jetre. A posebno ovo je fibrotest.
U algoritmu za dijagnosticiranje nealkoholne masne bolesti jetre ne obraćamo pažnju samo na specifične znakove, već i na aktivnost enzima alanin i asparaginske transaminaze, gama-glutamil transpeptidaze, alkalne fosfataze, a pazimo i na unos alkohola, o čemu su govorili prethodni kolege. I želio bih skrenuti pozornost, naravno, na faktore rizika: metabolički sindrom, inzulinska rezistencija, dijabetes. Propisuje se terapija za ispravljanje ove situacije, a po potrebi i biopsija jetre. Bez sumnje, potrebne su apsolutne indikacije za biopsiju. A ako indeks tjelesne mase prelazi 35 i 40, tada se već poduzimaju mjere koje se odnose na kirurško liječenje.
Skrećem pozornost na brojne lijekove (nesteroidne - protuupalne glukokortikoze i steroidne lijekove, tetraciklinske antibiotike), niz prehrambenih čimbenika (post, naglo mršavljenje, kirurški zahvati, metabolički genetski čimbenici, u posebno, nasljedna hemokromatoza, razni otrovi) i druge popratne bolesti. Ovo je vrlo važno za diferencijalnu dijagnozu.
U fazi steatoze važno je liječenje pretilosti, inzulinske rezistencije i dislipidemije. U fazi steatohepatitisa najvažniji trenutak je uklanjanje oksidativnog stresa, upale i fibroze.
Prekomjerna indukcija citokroma P450 2E ima štetne učinke na hepatocite zbog oslobađanja slobodnih radikala. Esencijalni fosfolipidi djeluju ne samo kao antioksidansi, već služe i kao vrlo važan čimbenik za smanjenje aktivnosti citokroma 2E1, kako je prikazano u radovima M. Aleynika. Rezultati nekih studija sugeriraju da uvođenje esencijalnih fosfolipida može smanjiti indukciju citokroma P450 2E (rad Vladimira Trofimoviča Ivaškina, koji je predstavljen s Marinom Viktorovnom Mayevskaya u ruskim izvorima 2004. godine).
Zvjezdaste stanice sudjeluju u nastanku završne faze nealkoholne masne bolesti jetre. A u laboratorijskim pokusima dokazano je da potpuna prevencija aktivacije zvjezdastih stanica pomoću inhibitora CYP2E1 sprječava razvoj ciroze.
Skrećem pozornost da ne samo ruski autor M. Aleynik, već i japanski autor Akiyama u časopisu “Hepatology” 2009. godine, na temelju modela alkoholnog oštećenja jetre, također obraća pažnju na citokrom P450. 2E, acetil-CoA oksidaze i nikotinamid adenin dinukleotid oksidaze, da esencijalni fosfolipidi pokazuju protuupalno, antiapoptotsko i antifibrotično djelovanje u ovoj patologiji.
Ovo je teoretska verzija pretpostavke o upotrebi inhibitora citokroma P450, a posebno lijeka "Essentiale", koji je referentna i najvažnija točka za inhibiciju citokroma P450 2E i, sukladno tome, P450 4A11. Time se sprječava oksidacija lipida, glikoliza i smanjuje sinteza masnih kiselina.
U liječenju nealkoholne masne bolesti jetre koriste se sljedeći lijekovi: inzulinski senzibilizatori, antioksidansi, hepatoprotektori, antimikrobici.
Ali želio bih skrenuti pozornost na membranske fosfolipide. Oni su glavne lipidne komponente staničnih membrana. Oštećenje fosfolipidnih membrana dovodi do sindroma citolize, a višak reaktivnih kisikovih vrsta dovodi do oštećenja fosfolipidnih membrana na temelju mikrosomalne γ-oksidacije i peroksimalne β-oksidacije. Sukladno tome, oštećenje fosfolipidnih membrana rezultira staničnim odumiranjem, što dovodi do inicijacije fibroze i aktivacije zvjezdastih stanica.
Oštećenje strukture jetre je oštećenje membrana. U verziji esencijalnih fosfolipida, radi se o materijalu koji obnavlja stanične membrane umjesto lipida. Obnavljanje strukture jetre omogućuje vraćanje funkcije jetre.
Naši pacijenti ne boluju samo od alkoholne masne bolesti jetre, alkoholnog hepatitisa, nego i od drugih bolesti jetre, to je nepobitna činjenica. Skrećem pozornost da prema E. Kunz (monografija iz 2008.) esencijalni fosfolipidi imaju antifibrotski učinak, učinak koji stabilizira žuč i membranu hepatocita.
Ovo je publikacija koja je objavljena 2008. godine na temelju farmakoloških i kliničkih podataka. Terapija esencijalnim fosfolipidima čini se preferiranim izborom za značajno smanjenje manifestacija i otklanjanje masne bolesti jetre različite etiologije, koja je nastala zbog konzumiranja alkohola, pretilosti, pa čak i ako se ne može utvrditi uzrok.
Želio bih istaknuti da postoji nekoliko studija o Essentialu. Ove studije su svima dobro poznate. Ali želio bih reći da čak i kod dijabetes melitusa Essentiale omogućuje pacijentima s nealkoholna bolest jetre za normalizaciju razine glukoze, glikiranog hemoglobina, serumskog kolesterola.
Zaključno, želio bih reći da je oštećenje jetre karakterizirano nakupljanjem masti u odsutnosti zlouporabe alkohola poznato kao nealkoholna masna bolest jetre. Čimbenici rizika uključuju pretilost i dijabetes tipa 2. U patogenezi nealkoholne masne bolesti jetre poseban značaj ima prekomjerna aktivnost citokroma P450 2E1. Kliničke varijante tijeka bolesti: bol u desnom hipohondriju, astenovegetativni i dispeptički poremećaji, hepatomegalija. A naš dijagnostički algoritam temelji se na dosljednom isključivanju alkoholnog i jatrogenog, kao i virusnog oštećenja jetre.
Citokrom P450(CYP450) velika je skupina enzima odgovornih za metabolizam stranih organskih spojeva i lijekova. Enzimi obitelji citokroma P450 provode oksidativnu biotransformaciju lijekova i niza drugih endogenih bioorganskih tvari te na taj način obavljaju funkciju detoksikacije. Citokromi su uključeni u metabolizam mnogih klasa lijekova, kao što su inhibitori protonske pumpe, antihistaminici, inhibitori retrovirusne proteaze, benzodiazepini, blokatori kalcijevih kanala i drugi.
Citokrom P450 je proteinski kompleks s kovalentno vezanim hemom (metaloprotein), koji osigurava dodavanje kisika. Hem je pak kompleks protoporfirina IX i dvovalentnog atoma željeza. Broj 450 označava da reducirani hem povezan s CO ima maksimalnu apsorpciju svjetlosti na valnoj duljini od 450 nm.
Citokromi P-450 sudjeluju ne samo u metabolizmu lijekova, već iu pretvorbi hemoglobina u bilirubin, sintezi steroida itd. Svi izoformi citokroma P-450 grupirani su u obitelji CYP1, CYP2, CYP3. Unutar obitelji razlikuju se potporodice A, B, C, D, E. Unutar potfamilija izoforme su označene serijskim brojem. Na primjer, CYP2C19 je naziv 19. po redu citokroma potfamilije "C", familije "2". Ukupno ih je oko 250 različite vrste citokroma P-450, od kojih se oko 50 nalazi u ljudskom tijelu, a samo šest (CYP1A2, CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4) povezano je s metabolizmom lijekova.
Na aktivnost citokroma P-450 utječu mnogi čimbenici – pušenje, alkohol, dob, genetika, prehrana, bolesti. Ovi čimbenici odgovorni su za formiranje individualnih karakteristika rada enzima P-450 i određuju učinke interakcije lijekova za konkretnog pacijenta.
Značaj citokroma P450 za gastroenterologiju
Nedavno povećani interes gastroenterologa za izoforme citokroma P450 CYP2C19 i CYP3A4 posljedica je njihove uloge u metabolizmu derivata benzimidazola, u koje spadaju svi lijekovi iz ATC skupine A02BC „Inhibitori protonske pumpe” (omeprazol, pantorazol, lansoprazol, rabeprazol i esomeprazol). ) . Klinički je značajno da je gen CYP2C19 polimorfan i vrijednost od terapeutski učinak razni PPI.Među PPI, lansoprazol pokazuje najveći inhibicijski učinak na CYP2C19, a slijede ga omeprazol i esomeprazol u manjoj mjeri. Učinak rabeprazola još je manji, ali njegov tioester, nastao tijekom neenzimskog metabolizma, ima značajan inhibicijski učinak na aktivnost CYP2C19. Pantoprazol ima najmanji učinak na CYP2C19. Pantoprazol ima najveći inhibitorni učinak na CYP3A4 in vitro, a slijede ga (kako se učinak smanjuje) omeprazol, esomeprazol, rabeprazol i lansoprazol. Za pacijente koji primaju više lijekova, pantoprazol je poželjniji među PPI (Bordin D.S.). 
Metabolizam pet inhibitora protonske pumpe.
Tamnije strelice označavaju značajnije metaboličke putove.
Slika preuzeta iz članka Marelli S., Pace F.
Uz aktivno sudjelovanje CYP3A4 dolazi do metabolizma domperidona, cisaprida i velikog broja drugih lijekova.
Brojni gastroenterološki lijekovi inhibiraju citokrom CYP3A4, čime utječu na farmakokinetiku lijekova koji se uzimaju zajedno.
Problem interakcije lijekova
U suvremenoj kliničkoj praksi, kombinirana uporaba lijekova je široko rasprostranjena, što je povezano s prisutnošću nekoliko bolesti u pacijenta ili nedovoljnom učinkovitošću monoterapije. Kod kombinirane terapije moguće su interakcije lijekova. Otprilike 56% pacijenata mlađih od 65 godina i 73% pacijenata starijih od 65 godina uzima više od jednog lijeka. Uzimanje dva lijeka dovodi do njihove interakcije u 6% bolesnika. Propisivanje 5 (ili 10) lijekova povećava stopu interakcija do 50 (ili 100)%.Potencijalno opasne kombinacije lijekovi su ozbiljan klinički problem. Postoje dokazi da je od 17 do 23% kombinacija lijekova koje propisuju liječnici potencijalno opasno. Samo u Sjedinjenim Američkim Državama, 48 tisuća pacijenata umre svake godine zbog neželjene interakcije lijekova. FDA je poništila registraciju nekoliko lijekova (uključujući prokinetik cisaprid) zbog njihovih potencijalno opasnih interakcija s drugim lijekovima, uključujući i smrtne ishode.
Glavni mehanizmi interakcija lijekova povezani su s promjenama njihove farmakokinetike ili farmakodinamike. Najznačajnije, prema suvremenim konceptima, su promjene u farmakokinetici tijekom metabolizma lijeka uz sudjelovanje citokroma P-450.
Primjer opasne interakcije je nedavno otkrivena interakcija IPP-a i klopidogrela, naširoko korištena u liječenju bolesnika s koronarna bolest srca. Kako bi se smanjio rizik od gastrointestinalnih komplikacija kod pacijenata koji primaju acetilsalicilna kiselina u kombinaciji s klopidogrelom propisuje se PPI. Budući da se bioaktivacija klopidogrela odvija uz sudjelovanje CYP2C19, uzimanje PPI-a koje metabolizira ovaj citokrom može smanjiti aktivaciju i antitrombocitni učinak klopidogrela. U svibnju 2009. na konferenciji Društva za kardiovaskularnu angiografiju i intervencije (SCAI) predstavljeni su podaci koji pokazuju da istodobna primjena klopidogrela i IPP-a značajno povećava rizik od infarkta miokarda, moždanog udara, nestabilne angine, potrebe za ponavljanjem koronarnih intervencija i koronarne smrti (Bordin D .S.).
Citokrom CYP2C19
Izoforma citokroma P450 CYP2C19 (S-mefenitoin hidroksilaza) katalizira reakcije 5-hidroksilacije piridinskog prstena i 5"-demetilacije benzimidazolskog prstena. U ljudskom tijelu CYP2C19 nalazi se u hepatocitima.Sve vrste mutacija gena CYP2C19 mogu se podijeliti u tri skupine:
- Bez mutacija (homozigoti), također su brzi metabolizatori PPI.
- Imajući mutaciju u jednom alelu (heterozigoti), srednji tip metabolizma.
- Imajući mutacije u oba alela, oni su također spori metabolizatori PPI-a.
| CYP2C19 genotip |
Prevalencija (Tkach S.M. i sur., 2006.) |
Vrsta metabolizma |
PPI poluživot, T½, sat (Lapina T.L.) |
Inhibicijski učinak PPI na kiselinu | ||
| bijele rase | Mongoloidna rasa | |||||
|
Nema mutacija (homozigoti) |
90% bijelaca | 50,6 % |
34,0 % |
Brzo | 1 | Kratak |
|
Mutacija u prvom redu (heterozigoti) |
10% bijelaca | 40,5 % | 47,6 % | Srednji | - | Prosjek |
| Mutacija u obje uličice | 20-30% azijskog stanovništva | 3,3 % | 18,4 % | Usporiti |
2–10 |
visoko |
Spori metabolizatori razlikuju se od brzih i srednjih metabolizatora dvostruko većom koncentracijom PPI u krvnoj plazmi i poluživotom. Polimorfizam gena koji kodira izoformu 2C19 određuje različite brzine metabolizma PPI u bolesnika. U vezi s navedenim, odabir PPI-a preporučuje se provoditi pod nadzorom dnevna pH-metrija(Khavkin A.I., Zhikhareva N.S., Drozdovskaya N.V.).
- CYP2C19 aktivno metabolizira sljedeće lijekove: tricikličke antidepresive (amitriptilin, klomipramin, imipramin), antidepresive - selektivni inhibitor ponovne pohrane serotonina citalopram, antidepresiv - MAO inhibitor moklobemid, antikonvulzive i antiepileptike (diazepam, primidon, fenitoin, fenobarbital, nordazepam), pro ton-konvulzivan inhibitori MP (omeprazol, pantorazol, lansoprazol, rabeprazol i esomeprazol), lijek protiv malarije progvanil, nesteroidni protuupalni lijekovi diklofenak i indometacin, kao i: varfarin, gliklazid, klopidogrel, propranolol, ciklofosfamid, nelfinavir, progesteron, tenipozid, tetrahidrokanabinol, karisopro dol, vorikonazol i drugi
- jaki inhibitori CYP2C19: moklobemid, fluvoksamin, kloramfenikol (kloramfenikol)
- nespecifični inhibitori CYP2C19: PPI omeprazol i lansoprazol, H2-blokator cimetidin, NSAID indometacin, kao i fluoksetin, felbamat, ketokonazol, modafinil, okskarbazepin, probenecid, tiklopidin, topiramat
- Induktori CYP2C19: rifampicin, artemisinin, karbamazepin, noretisteron, prednizon, gospina trava.
Učinak različitih genotipova CYP2C19 na učinkovitost eradikacije Helicobacter pylori
Bolesnici s genotipom "brzih" metabolizatora imaju brz metabolizam inhibitora protonske pumpe, stoga je antisekretorni učinak uzimanja potonjeg kod njih manje izražen nego kod osoba s fenotipovima "srednjih" i "sporih" metabolizatora. Razlika u antisekretornom učinku može odrediti više niska razina iskorjenjivanje Helicobacter pylori kod “brzih” metabolizatora. Dakle, postoji više visoka efikasnost eradikacijska terapija u bolesnika s genotipovima „sporih” (88,9%) i „srednjih” (82,7%) metabolizatora veća je u usporedbi s „brzim” (vidi sliku). 
Utjecaj različitih genotipova CYP2C19 na učinkovitost eradikacije Helicobacter pylori.
BM – “brzi” metabolizatori, PM – “srednji” metabolizatori, MM – “spori” metabolizatori (Maev I.V. et al.)
Zbog činjenice da su molekularno-genetske studije nedostupne liječniku, na "brze" metabolizatore se može posumnjati na temelju postojanosti bolnog sindroma u trbuhu 3-4 dana od početka uzimanja PPI-a, kao i uzimajući u obzir spora endoskopska dinamika tijekom epitelizacije erozija i ožiljnih ulcerativnih defekata u bolesnika. S druge strane, nedostatnost antisekretornog učinka IPP terapije može se potvrditi metodom dnevne intragastrične pH-metrije (Maev I.V. et al.).
Citokrom CYP3A4
Enzim CYP3A4 katalizira reakciju sulfoksidacije, što dovodi do stvaranja sulfonske skupine. CYP3A4 jedan je od najvažnijih citokroma za lijekove, budući da biotransformira, barem djelomično, oko 60% oksidiranih lijekova. Iako aktivnost CYP3A4 uvelike varira, nije podložna genetskom polimorfizmu. Položaj CYP3A4 na apikalnim membranama enterocita i hepatocita tankog crijeva olakšava metabolizam lijekova prije ulaska lijeka u sistemsku cirkulaciju, što je poznato kao "učinak prvog prolaza".Genetski defekt u CYP3A4 može uzrokovati razvoj sekundarnog dugotrajnog sindroma. QT interval kod uzimanja cisaprida i, kao posljedica toga, razvoj srčane aritmije (Khavkin A.I. et al.).
- CYP3A4 je glavni enzim u metabolizmu sljedećih lijekova: imunosupresivi (ciklosporin, sirolimus, takrolimus), lijekovi koji se koriste u kemoterapiji (anastrozol, ciklofosfamid, docetaksel, erlotinib, tirfostin, etopozid, ifosfamid, paklitaksel, tamoksifen, tenipozid, vinblastin, vindesin , gefitinib), antifungici (klotrimazol, ketokonazol, itrakonazol),
Citokrom P450(CYP450) velika je skupina enzima odgovornih za metabolizam stranih organskih spojeva i lijekova. Enzimi obitelji citokroma P450 provode oksidativnu biotransformaciju lijekova i niza drugih endogenih bioorganskih tvari te na taj način obavljaju funkciju detoksikacije. Citokromi su uključeni u metabolizam mnogih klasa lijekova, kao što su inhibitori protonske pumpe, antihistaminici, inhibitori retrovirusne proteaze, benzodiazepini, blokatori kalcijevih kanala i drugi.
Citokrom P450 je proteinski kompleks s kovalentno vezanim hemom (metaloprotein), koji osigurava dodavanje kisika. Hem je pak kompleks protoporfirina IX i dvovalentnog atoma željeza. Broj 450 označava da reducirani hem povezan s CO ima maksimalnu apsorpciju svjetlosti na valnoj duljini od 450 nm.
Citokromi P-450 sudjeluju ne samo u metabolizmu lijekova, već iu pretvorbi hemoglobina u bilirubin, sintezi steroida itd. Svi izoformi citokroma P-450 grupirani su u obitelji CYP1, CYP2, CYP3. Unutar obitelji razlikuju se potporodice A, B, C, D, E. Unutar potfamilija izoforme su označene serijskim brojem. Na primjer, CYP2C19 je naziv 19. po redu citokroma potfamilije "C", familije "2". Ukupno postoji oko 250 različitih vrsta citokroma P-450, od kojih se oko 50 nalazi u ljudskom tijelu, a samo njih šest (CYP1A2, CYP2C9, CYP2C19, CYP2D6, CYP2E1, CYP3A4) relevantni su za metabolizam lijekova.
Na aktivnost citokroma P-450 utječu mnogi čimbenici – pušenje, alkohol, dob, genetika, prehrana, bolesti. Ovi čimbenici odgovorni su za formiranje individualnih karakteristika rada enzima P-450 i određuju učinke interakcija lijekova u određenom pacijentu.
Značaj citokroma P450 za gastroenterologiju
Nedavno povećani interes gastroenterologa za izoforme citokroma P450 CYP2C19 i CYP3A4 posljedica je njihove uloge u metabolizmu derivata benzimidazola, u koje spadaju svi lijekovi iz ATC skupine A02BC „Inhibitori protonske pumpe” (omeprazol, pantorazol, lansoprazol, rabeprazol i esomeprazol). ) . Klinički je značajno da je gen CYP2C19 polimorfan, a veličina terapijskog učinka različitih PPI uvelike ovisi o stanju ovog gena u bolesnika.Među PPI, lansoprazol pokazuje najveći inhibicijski učinak na CYP2C19, a slijede ga omeprazol i esomeprazol u manjoj mjeri. Učinak rabeprazola još je manji, ali njegov tioester, nastao tijekom neenzimskog metabolizma, ima značajan inhibicijski učinak na aktivnost CYP2C19. Pantoprazol ima najmanji učinak na CYP2C19. Pantoprazol ima najveći inhibitorni učinak na CYP3A4 in vitro, a slijede ga (kako se učinak smanjuje) omeprazol, esomeprazol, rabeprazol i lansoprazol. Za pacijente koji primaju više lijekova, pantoprazol je poželjniji među PPI (Bordin D.S.).
Metabolizam pet inhibitora protonske pumpe.
Tamnije strelice označavaju značajnije metaboličke putove.
Slika preuzeta iz članka Marelli S., Pace F.
Uz aktivno sudjelovanje CYP3A4 dolazi do metabolizma domperidona, cisaprida i velikog broja drugih lijekova.
Brojni gastroenterološki lijekovi inhibiraju citokrom CYP3A4, čime utječu na farmakokinetiku lijekova koji se uzimaju zajedno.
Problem interakcije lijekova
U suvremenoj kliničkoj praksi, kombinirana uporaba lijekova je široko rasprostranjena, što je povezano s prisutnošću nekoliko bolesti u pacijenta ili nedovoljnom učinkovitošću monoterapije. Kod kombinirane terapije moguće su interakcije lijekova. Otprilike 56% pacijenata mlađih od 65 godina i 73% pacijenata starijih od 65 godina uzima više od jednog lijeka. Uzimanje dva lijeka dovodi do njihove interakcije u 6% bolesnika. Propisivanje 5 (ili 10) lijekova povećava stopu interakcija do 50 (ili 100)%.Potencijalno opasne kombinacije lijekova ozbiljan su klinički problem. Postoje dokazi da je od 17 do 23% kombinacija lijekova koje propisuju liječnici potencijalno opasno. Samo u Sjedinjenim Američkim Državama, 48 tisuća pacijenata umre svake godine zbog neželjene interakcije lijekova. FDA je poništila registraciju nekoliko lijekova (uključujući prokinetik cisaprid) zbog njihovih potencijalno opasnih interakcija s drugim lijekovima, uključujući i smrtne ishode.
Glavni mehanizmi interakcija lijekova povezani su s promjenama njihove farmakokinetike ili farmakodinamike. Najznačajnije, prema suvremenim konceptima, su promjene u farmakokinetici tijekom metabolizma lijeka uz sudjelovanje citokroma P-450.
Primjer opasne interakcije je nedavno otkrivena interakcija između PPI i klopidogrela, koji se naširoko koristi u liječenju bolesnika s koronarnom bolesti srca. Kako bi se smanjio rizik od gastrointestinalnih komplikacija, bolesnicima koji primaju acetilsalicilnu kiselinu u kombinaciji s klopidogrelom propisuje se IPP. Budući da se bioaktivacija klopidogrela odvija uz sudjelovanje CYP2C19, uzimanje PPI-a koje metabolizira ovaj citokrom može smanjiti aktivaciju i antitrombocitni učinak klopidogrela. U svibnju 2009. na konferenciji Društva za kardiovaskularnu angiografiju i intervencije (SCAI) predstavljeni su podaci koji pokazuju da istodobna primjena klopidogrela i IPP-a značajno povećava rizik od infarkta miokarda, moždanog udara, nestabilne angine, potrebe za ponavljanjem koronarnih intervencija i koronarne smrti (Bordin D .S.).
Citokrom CYP2C19
Izoforma citokroma P450 CYP2C19 (S-mefenitoin hidroksilaza) katalizira reakcije 5-hidroksilacije piridinskog prstena i 5"-demetilacije benzimidazolskog prstena. U ljudskom tijelu CYP2C19 nalazi se u hepatocitima.Sve vrste mutacija gena CYP2C19 mogu se podijeliti u tri skupine:
- Bez mutacija (homozigoti), također su brzi metabolizatori PPI.
- Imajući mutaciju u jednom alelu (heterozigoti), srednji tip metabolizma.
- Imajući mutacije u oba alela, oni su također spori metabolizatori PPI-a.
| CYP2C19 genotip |
Prevalencija (Tkach S.M. i sur., 2006.) |
Vrsta metabolizma |
PPI poluživot, T½, sat (Lapina T.L.) |
Inhibicijski učinak PPI na kiselinu | ||
| bijele rase | Mongoloidna rasa | |||||
|
Nema mutacija (homozigoti) |
90% bijelaca | 50,6 % |
34,0 % |
Brzo | 1 | Kratak |
|
Mutacija u prvom redu (heterozigoti) |
10% bijelaca | 40,5 % | 47,6 % | Srednji | - | Prosjek |
| Mutacija u obje uličice | 20-30% azijskog stanovništva | 3,3 % | 18,4 % | Usporiti |
2–10 |
visoko |
Spori metabolizatori razlikuju se od brzih i srednjih metabolizatora dvostruko većom koncentracijom PPI u krvnoj plazmi i poluživotom. Polimorfizam gena koji kodira izoformu 2C19 određuje različite brzine metabolizma PPI u bolesnika. U vezi s navedenim, odabir PPI-a preporučuje se provoditi pod nadzorom dnevna pH-metrija(Khavkin A.I., Zhikhareva N.S., Drozdovskaya N.V.).
- CYP2C19 aktivno metabolizira sljedeće lijekove: tricikličke antidepresive (amitriptilin, klomipramin, imipramin), antidepresive - selektivni inhibitor ponovne pohrane serotonina citalopram, antidepresiv - MAO inhibitor moklobemid, antikonvulzive i antiepileptike (diazepam, primidon, fenitoin, fenobarbital, nordazepam), pro ton-konvulzivan inhibitori MP (omeprazol, pantorazol, lansoprazol, rabeprazol i esomeprazol), lijek protiv malarije progvanil, nesteroidni protuupalni lijekovi diklofenak i indometacin, kao i: varfarin, gliklazid, klopidogrel, propranolol, ciklofosfamid, nelfinavir, progesteron, tenipozid, tetrahidrokanabinol, karisopro dol, vorikonazol i drugi
- jaki inhibitori CYP2C19: moklobemid, fluvoksamin, kloramfenikol (kloramfenikol)
- nespecifični inhibitori CYP2C19: PPI omeprazol i lansoprazol, H2-blokator cimetidin, NSAID indometacin, kao i fluoksetin, felbamat, ketokonazol, modafinil, okskarbazepin, probenecid, tiklopidin, topiramat
- Induktori CYP2C19: rifampicin, artemisinin, karbamazepin, noretisteron, prednizon, gospina trava.
Utjecaj različitih genotipova CYP2C19 na učinkovitost eradikacije Helicobacter pylori
Bolesnici s genotipom "brzih" metabolizatora imaju brz metabolizam inhibitora protonske pumpe, stoga je antisekretorni učinak uzimanja potonjeg kod njih manje izražen nego kod osoba s fenotipovima "srednjih" i "sporih" metabolizatora. Razlike u antisekretornom učinku mogu odrediti niže stope eradikacije Helicobacter pylori kod “brzih” metabolizatora. Dakle, postoji veća učinkovitost eradikacijske terapije u bolesnika s genotipovima "sporih" (88,9%) i "srednjih" (82,7%) metabolizatora u usporedbi s "brzim" metabolizatorima (vidi sliku).
Utjecaj različitih genotipova CYP2C19 na učinkovitost eradikacije Helicobacter pylori.
BM – “brzi” metabolizatori, PM – “srednji” metabolizatori, MM – “spori” metabolizatori (Maev I.V. et al.)
Zbog činjenice da su molekularno-genetske studije nedostupne liječniku, na "brze" metabolizatore se može posumnjati na temelju postojanosti bolnog sindroma u trbuhu 3-4 dana od početka uzimanja PPI-a, kao i uzimajući u obzir spora endoskopska dinamika tijekom epitelizacije erozija i ožiljnih ulcerativnih defekata u bolesnika. S druge strane, nedostatnost antisekretornog učinka IPP terapije može se potvrditi metodom dnevne intragastrične pH-metrije (Maev I.V. et al.).
Citokrom CYP3A4
Enzim CYP3A4 katalizira reakciju sulfoksidacije, što dovodi do stvaranja sulfonske skupine. CYP3A4 jedan je od najvažnijih citokroma za lijekove, budući da biotransformira, barem djelomično, oko 60% oksidiranih lijekova. Iako aktivnost CYP3A4 uvelike varira, nije podložna genetskom polimorfizmu. Položaj CYP3A4 na apikalnim membranama enterocita i hepatocita tankog crijeva olakšava metabolizam lijekova prije ulaska lijeka u sistemsku cirkulaciju, što je poznato kao "učinak prvog prolaza".Genetski defekt u CYP3A4 može biti uzrok razvoja sekundarnog sindroma produljenog QT intervala tijekom uzimanja cisaprida i, kao posljedica toga, razvoja srčane aritmije (Khavkin A.I. et al.).
- CYP3A4 je glavni enzim u metabolizmu sljedećih lijekova: imunosupresivi (ciklosporin, sirolimus, takrolimus), lijekovi koji se koriste u kemoterapiji (anastrozol, ciklofosfamid, docetaksel, erlotinib, tirfostin, etopozid, ifosfamid, paklitaksel, tamoksifen, tenipozid, vinblastin, vindesin , gefitinib), antifungici (klotrimazol, ketokonazol, itrakonazol),
P450 su membranski proteini.
Sustav citokroma P450 uključen je u oksidaciju brojnih spojeva, endogenih i egzogenih. Enzimi ove skupine imaju važnu ulogu u metabolizmu steroida, žučnih kiselina, nezasićenih masnih kiselina, fenolnih metabolita, kao iu neutralizaciji ksenobiotika (lijekovi, otrovi, lijekovi).
Reakcije koje uključuju sustav citokroma P450
Monooksigenaze ovisne o citokromu P450 kataliziraju razgradnju razne tvari kroz hidroksilacija uz sudjelovanje donora elektrona NADP H i molekularnog kisika. U ovoj reakciji, jedan atom kisika se dodaje supstratu, a drugi se reducira u vodu.
Enzimi obitelji citokroma P450, za razliku od drugih hemoproteina, koji obično imaju jednu vrstu aktivnosti i strogo definiranu funkciju, vrlo su raznoliki u funkciji, tip enzimska aktivnost, često imaju nisku specifičnost supstrata. P450 mogu pokazivati i aktivnost monooksigenaze i oksigenaze i stoga se ponekad nazivaju oksidaze miješane funkcije.
Reakcije oksigenaze katalizirane citokromom P450 vrlo su raznolike. Jedna od najčešćih reakcija oksidacije ksenobiotika je oksidativna dealkilacija, praćena oksidacijom alkilne skupine vezane na atome N, O ili S. Taj se proces odvija u endoplazmatskom retikulumu (ER) hepatocita. Njihova supstratna specifičnost je niska. Oni najučinkovitije kataliziraju oksidaciju nepolarnih spojeva s alifatskim ili aromatskim prstenovima. Jetreni P450 je, između ostalog, uključen u oksidaciju alkohola u odgovarajuće aldehide. Hidroksilacija hidrofobnih spojeva poboljšava njihovu topljivost u vodi i potiče izlučivanje putem bubrega. U razliciti ljudi skup citokroma P450 u ER-u razlikuje se zbog genetskih karakteristika. U tom smislu, proučavanje enzimatskog sustava P450 od velike je važnosti za farmakologiju. Svi ostali enzimi iz obitelji P450 lokalizirani su na * , a njihovi katalitički centri okrenuti su prema matrici.
Drugi uobičajeni tip reakcije je hidroksilacija cikličkih spojeva (aromatskih, zasićenih i heterocikličkih ugljikovodika). Enzimi obitelji P450 također mogu katalizirati reakcije hidroksilacije alifatskih spojeva, N-oksidaciju, oksidativnu deaminaciju i reakcije redukcije nitro spojeva.
Geni ljudskog citokroma P450
| Obitelj | Funkcije | Spoj | Naslovi |
| CYP1 | metabolizam lijekova i steroida (osobito estrogena) | 3 podfamilije, 3 gena, 1 pseudogen | CYP1A1, CYP1A2, CYP1B1 |
| CYP2 | metabolizam lijekova i steroida | 13 podfamilija, 16 gena, 16 pseudogena | CYP2A6, CYP2A7, CYP2A13, CYP2B6, CYP2C8, CYP2C9, CYP2C18, CYP2C19, CYP2D6, CYP2E1, CYP2F1, CYP2J2, CYP2R1, CYP2S1, CYP2U1, CYP2W1 |
| CYP3 | metabolizam lijekova i steroida (uključujući testosteron) | 1 podfamilija, 4 gena, 2 pseudogena | CYP3A4, CYP3A5, CYP3A7, CYP3A43 |
| CYP4 | metabolizam arahidonske kiseline | 6 podfamilija, 12 gena, 10 pseudogena | CYP4A11, CYP4A22, CYP4B1, CYP4F2, CYP4F3, CYP4F8, CYP4F11, CYP4F12, CYP4F22, CYP4V2, CYP4X1, CYP4Z1 |
| CYP5 | sinteza tromboksana A 2 | 1 podfamilija, 1 gen | CYP5A1 (tromboksan A 2 sintaza) |
| CYP7 | biosinteza žučnih kiselina, sudjelovanje u metabolizmu steroida | 2 podfamilije, 2 gena | CYP7A1, CYP7B1 |
| CYP8 | razne | 2 podfamilije, 2 gena | CYP8A1 (sinteza prostaciklina), CYP8B1 (biosinteza žučne kiseline) |
| CYP11 | biosinteza steroida | 2 podfamilije, 3 gena | CYP11A1, CYP11B1, CYP11B2 |
| CYP17 | biosinteza steroida, 17-alfa hidroksilaza | 1 podfamilija, 1 gen | CYP17A1 |
| CYP19 | biosinteza steroida (aromataza, koja sintetizira estrogen) | 1 podfamilija, 1 gen | CYP19A1 |
| CYP20 | nije instalirano | 1 podfamilija, 1 gen | CYP20A1 |
| CYP21 | biosinteza steroida | 2 podfamilije, 1 gen, 1 pseudogen | CYP21A2 |
| CYP24 | biorazgradnja vitamina D | 1 podfamilija, 1 gen | CYP24A1 |
| CYP26 | hidroksilacija retinolne kiseline | 3 podfamilije, 3 gena | CYP26A1, CYP26B1, CYP26C1 |
| CYP27 | razne | 3 podfamilije, 3 gena | CYP27A1 (biosinteza žučnih kiselina), CYP27B1 (vitamin D 3 1-alfa-hidroksilaza, aktiviranje vitamina D 3), CYP27C1 (funkcija nepoznata) |
| CYP39 | 7-alfa-hidroksilacija 24-hidroksikolesterola | 1 podfamilija, 1 gen | CYP39A1 |
| CYP46 | kolesterol 24-hidroksilaza | 1 podfamilija, 1 gen | CYP46A1 |
| CYP51 | biosinteza kolesterola | 1 podfamilija, 1 gen, 3 pseudogena | CYP51A1 (14 alfa demetilaza lanosterol) |
Napišite recenziju o članku "Citokrom P450"
Bilješke
- , sa. 180-181 (prikaz, stručni).
- , sa. 310-311 (prikaz, ostalo).
- Danielson P. B.(engleski) // Current drug metabolism. - 2002. - Vol. 3, br. 6. - Str. 561-597. - PMID 12369887.ispraviti
- Ortiz de Montellano, Paul R. Citokrom P450: struktura, mehanizam i biokemija. - 3. izdanje. - New York: Kluwer Academic/Plenum Publishers, 2005. - ISBN 0-306-48324-6.
- , sa. 348-349 (prikaz, ostalo).
- .
Književnost
- D. Nelson, M. Cox. Osnove Lehningerove biokemije: u 3 sveska - M.: BINOM, 2014. - T. 2. - P. 348-349. - 636 s. - ISBN 978-5-94774-366-1.
- Britton G.. - Moskva: Mir, 1986. - 422 str. - 3050 primjeraka.
- Jan Kolman, Klaus-Heinrich Rehm.= Taschenatlas der Biochemie. - Moskva: Mir, 2000. - 470 str. - 7000 primjeraka.
- Ponomarenko T. M., Sychev D. A., Chikalo A. O., Berdnikova N. G., Kukes V. G.// Farmakokinetika i farmakodinamika. - 2012. - br.1. - str. 25-28.
Linkovi
- .
- .
- .
Izvadak koji karakterizira citokrom P450
I grofica i Sonya razumjele su da Moskva, vatra Moskve, što god ona bila, naravno, ne može biti važna Natashi.Grof je opet otišao iza pregrade i legao. Grofica priđe Nataši, dotakne joj glavu naopako, kao kad joj je kći bila bolesna, zatim joj dotakne usnama čelo, kao da želi vidjeti ima li groznica, i poljubi je.
-Hladno ti je. Treseš se cijelim tijelom. Trebao bi otići u krevet”, rekla je.
- Idi u krevet? Da, dobro, idem u krevet. Idem sad u krevet, rekla je Natasha.
Budući da je Nataši jutros rečeno da je princ Andrej ozbiljno ranjen i da ide s njima, tek u prvoj minuti je mnogo pitala gdje? Kako? Je li opasno ozlijeđen? i smije li ga vidjeti? No, nakon što joj je rečeno da ga ne može vidjeti, da je teško ranjen, ali da nije životno ugrožen, ona, očito, nije vjerovala u ono što joj je rečeno, već je bila uvjerena da koliko god govorila, odgovorila bi isto, prestala pitati i govoriti. Cijelim putem, krupnih očiju, koje je grofica tako dobro poznavala i čijeg se izraza grofica tako bojala, Nataša je nepomično sjedila u kutu kočije i sada je tako sjedila na klupi na koju je sjela. O nečem je razmišljala, o nečemu je odlučivala ili je već odlučila u mislima sada - to je grofica znala, ali što je, nije znala, i to ju je plašilo i mučilo.
- Nataša, skini se, draga, lezi na moj krevet. (Samo je grofica sama dala napraviti krevet na krevetu; m ja Schoss i obje mlade dame morali smo spavati na podu na sijenu.)
„Ne, mama, leći ću ovde na pod“, rekla je Nataša ljutito, prišla prozoru i otvorila ga. Jasnije se čulo ađutantovo stenjanje s otvorenog prozora. Promolila je glavu u vlažni noćni zrak, a grofica je vidjela kako joj se mršava ramena tresu od jecaja i udaraju o tijelo. Nataša je znala da princ Andrej nije taj koji ječi. Znala je da princ Andrej leži u istoj vezi gdje su bili i oni, u drugoj kolibi s druge strane hodnika; ali ju je ovaj strašni neprestani jecaj natjerao na jecanje. Grofica je razmijenila poglede sa Sonyom.
- Lezi, draga moja, lezi, prijatelju - reče grofica, blago dodirujući rukom Natashino rame. - Pa, idi u krevet.
“Oh, da... Idem sad u krevet”, rekla je Natasha žurno se svlačeći i trgajući uzice sa suknje. Skinuvši haljinu i obukavši sako, podvila je noge, sjela na krevet pripremljen na podu i, prebacivši kratku tanku pletenicu preko ramena, počela je plesti. Tanki, dugi, poznati prsti brzo, spretno rastavljaju, pletu i vezuju pletenicu. Natashina glava okrenula se uobičajenim pokretom, prvo u jednom, zatim u drugom smjeru, ali su joj oči, grozničavo otvorene, gledale ravno i nepomično. Kad je noćno odijelo bilo gotovo, Natasha se tiho spustila na plahtu položenu na sijeno na rubu vrata.
"Natasha, lezi u sredinu", rekla je Sonya.
"Ne, tu sam", rekla je Natasha. "Idi u krevet", dodala je ljutito. I zarila je lice u jastuk.
Grofica, m me Schoss i Sonya žurno su se svukle i legle. U sobi je ostala jedna lampa. Ali u dvorištu je bilo sve svjetlije od požara Malih Mitišća, udaljenih dvije milje, i pijani povici naroda brujali su u krčmi, koju su Mamonovi kozaci razbili, na raskršću, na ulici, i neprestani jauk ađutanta se još moglo čuti.
Natasha je dugo slušala unutarnje i vanjske zvukove koji su dopirali do nje i nije se pomaknula. Najprije je čula molitvu i uzdahe svoje majke, pucketanje njezina kreveta ispod nje, poznato zviždanje hrkanja m me Schossa, tiho Sonyino disanje. Tada je grofica pozvala Natashu. Natasha joj nije odgovorila.
"Čini se da spava, mama", tiho je odgovorila Sonya. Grofica je, nakon što je neko vrijeme šutjela, opet zazvala, ali joj nitko nije odgovorio.
Ubrzo nakon toga, Natasha je čula majčino ravnomjerno disanje. Natasha se nije pomaknula, unatoč činjenici da joj je malo boso stopalo, nakon što je pobjeglo ispod pokrivača, bilo hladno na golom podu.
Kao da slavi pobjedu nad svima, u pukotini je vrisnuo cvrčak. Pijetao je daleko zapjevao, a voljeni su se odazvali. U krčmi je utihnula vriska, čulo se samo stajanje istog ađutanta. Natasha je ustala.
- Sonya? spavaš li? Majka? – šapnula je. Nitko se nije javio. Nataša je polako i pažljivo ustala, prekrižila se i oprezno zakoračila svojom uskom i gipkom bosom nogom na prljav, hladan pod. Podnica je zaškripala. Ona je, brzo mičući nogama, pretrčala nekoliko koraka poput mačića i zgrabila hladan nosač vrata.
Činilo joj se da nešto teško, ravnomjerno udarajući, kuca o sve zidove kolibe: to je njezino srce, zaleđeno od straha, od užasa i ljubavi, tuklo, pucalo.
Otvorila je vrata, prešla prag i zakoračila na vlažno, hladno tlo hodnika. Hladna hladnoća ju je osvježila. Opipala je usnulog čovjeka bosom nogom, opkoračila ga i otvorila vrata kolibe u kojoj je ležao princ Andrej. U ovoj je kolibi bilo mračno. U stražnjem kutu kreveta, na kojemu je nešto ležalo, na klupi je stajala svijeća od loja koja je dogorjela poput velike gljive.
Nataša je ujutro, kada su joj rekli za ranu i prisutnost princa Andreja, odlučila da ga treba vidjeti. Nije znala čemu, ali je znala da će susret biti bolan, a bila je još uvjerenija da je potreban.
Cijeli je dan živjela samo u nadi da će ga noću vidjeti. Ali sada, kada je došao ovaj trenutak, obuzeo ju je užas onoga što će vidjeti. Kako je osakaćen? Što je od njega ostalo? Je li bio poput onog neprestanog stenjanja ađutanta? Da, bio je takav. On je u njezinoj mašti bio personifikacija tog strašnog jecaja. Kad je ugledala nejasnu masu u kutu i zamijenila njegova podignuta koljena pod pokrivačem za njegova ramena, zamislila je nekakvo užasno tijelo i zastala užasnuta. Ali neodoljiva sila vukla ju je naprijed. Oprezno je napravila jedan korak, pa drugi i našla se usred male, pretrpane kolibe. U kolibi, ispod ikona, još jedna osoba ležala je na klupama (bio je to Timokhin), a još dvije osobe ležale su na podu (to su bili liječnik i sobar).
Sobar je ustao i nešto šapnuo. Timokhin, koji je patio od bolova u ranjenoj nozi, nije spavao i svim je očima gledao neobičnu pojavu djevojke u jadnoj košulji, sakou i vječnoj kapi. Pospane i preplašene riječi sobara; “Što ti treba, zašto?” - samo su prisilili Natashu da brzo priđe onome što je ležalo u kutu. Bez obzira na to koliko je ovo tijelo strašno ili neslično ljudskom, morala ga je vidjeti. Prošla je pored sobara: spaljena gljiva svijeće je otpala i jasno je vidjela princa Andreja kako leži ispruženih ruku na pokrivaču, baš kao što ga je uvijek vidjela.
Bio je isti kao i uvijek; ali užarena boja njegova lica, njegove svjetlucave oči, oduševljeno uprte u nju, a osobito nježan dječji vrat koji je virio iz sklopljene kragne njegove košulje, davali su mu poseban, nevin, dječji izgled, kakav ona, međutim, nikada nije vidjela u knezu Andreju. Prišla mu je i brzim, gipkim, mladenačkim pokretom kleknula.
Nasmiješio se i pružio joj ruku.
Za princa Andreja prošlo je sedam dana otkako se probudio u previjalištu Borodinskog polja. Cijelo to vrijeme bio je u gotovo stalnoj nesvijesti. Groznica i upala crijeva, koja su bila oštećena, po mišljenju liječnika koji je putovao s ranjenikom, trebala ga je odnijeti. Ali sedmoga dana s veseljem je pojeo krišku kruha s čajem, a liječnik je primijetio da se opća groznica smanjila. Knez Andrej se ujutro osvijestio. Prve noći nakon odlaska iz Moskve bilo je prilično toplo, a princ Andrej je ostao prenoćiti u kočiji; ali u Mytishchiju je sam ranjenik zahtijevao da ga iznesu i da mu daju čaj. Od boli koju je prouzročio unošenje u kolibu princ Andrej je glasno zastenjao i ponovno izgubio svijest. Kada su ga položili na kamperski ležaj, dugo je ležao zatvorenih očiju bez micanja. Zatim ih je otvorio i tiho šapnuo: “Što da imam za čaj?” Ovo sjećanje na male detalje života zadivilo je liječnika. Opipao je puls i, na svoje iznenađenje i nezadovoljstvo, primijetio da je puls bolji. Na njegovo nezadovoljstvo, liječnik je to primijetio jer je iz svog iskustva bio uvjeren da knez Andrej ne može živjeti i da će, ako sada ne umre, umrijeti uz velike patnje nešto kasnije. S knezom Andrejem nosili su bojnika njegove pukovnije Timohina, koji im se pridružio u Moskvi s crvenim nosom i bio ranjen u nogu u istoj bitki kod Borodina. S njima su jahali liječnik, prinčev sobar, njegov kočijaš i dva bolničara.
Princ Andrej je dobio čaj. Pohlepno je pio, grozničavim očima gledajući ispred sebe u vrata, kao da pokušava nešto shvatiti i zapamtiti.
- Ne želim više. Je li Timokhin ovdje? - upitao. Timohin je puzao prema njemu duž klupe.
- Ovdje sam, vaša ekselencijo.
- Kako je rana?
- Moje onda? Ništa. Jesi li to ti? “Princ Andrej je ponovno počeo razmišljati, kao da se nečega prisjeća.
-Mogu li dobiti knjigu? - On je rekao.
- Koja knjiga?
- Gospel! Ja nemam.
Liječnik je obećao da će ga dobiti i počeo ispitivati princa kako se osjeća. Knez Andrej je nevoljko, ali mudro odgovorio na sva pitanja liječnika, a zatim rekao da mu treba staviti jastuk, inače bi bilo neugodno i vrlo bolno. Liječnik i sobar podigoše kaput kojim je bio pokriven i, trznuvši se od teškog zadaha pokvarenog mesa koji se širio iz rane, počeše pregledavati ovo strašno mjesto. Liječnik je bio nečim jako nezadovoljan, nešto je promijenio, preokrenuo ranjenika tako da je on opet zastenjao i od boli pri okretanju opet izgubio svijest i počeo buncati. Stalno je govorio kako će mu što prije nabaviti ovu knjigu i staviti je tamo.
Citokrom P450. Autori otkrića, M. Klingerberg i D. Garfinkel, ustanovili su da se ovaj enzim, na temelju kemijske prirode prostetske skupine, može klasificirati kao citokrome tipa i. T. Omura i R. Sato otkrili su 1964. da kompleks reduciranog hemoproteina s ugljikovim monoksidom ima karakterističan maksimum na 450 nm, što je odredilo naziv enzima. Međutim, korištenje riječi "citokrom" u odnosu na hemoproteine klase P450 ne može se smatrati uspješnim, budući da je funkcija citokroma prijenos elektrona, a ne kataliza monooksigenaznih reakcija. U preporukama o nomenklaturi obitelji P450 koju je predložio D. Nebertomb, riječ "citokrom" spominje se samo pri dešifriranju oznake CYP (tj. citokroma Z450), koja se koristi za označavanje gena P450.
Trenutno je poznato više od 150 različitih P450, pronađenih u životinjama, biljkama, gljivama i bakterijama. Samo striktno anaerobne bakterije nemaju hemoprotein. Prokarioti sadrže topljivi P450. Prijelaz na eukariotske sustave prati ugradnja P450 u membranu, kao u slučaju kvasca i gljivica. Svi citokromi P450 viših organizama su membranski enzimi. U evolucijskom smislu, najstarija je bakterijska monooksigenaza
Na srednjem stupnju evolucijske ljestvice nalazi se mitohondrijski hidroksilazni sustav nadbubrežnih žlijezda. Ima sve značajke bakterijski topljivog sustava i također se sastoji od tri komponente. Dvije njegove komponente - flavoprotein koji sadrži FAD (NADPH- ili NADH-ovisna reduktaza) i ne-hemski protein koji sadrži sumpor (adrenodoksin) - topljivi su u vodi i lokalizirani u mitohondrijskom matriksu, a treća - P450 ugrađena je u membranu . Zanimljiva je visoka supstratna specifičnost mitohondrijskih hemoproteina, što ovaj sustav čini još sličnijim bakterijskom. Mitohondrijski citokromi P450 uglavnom su uključeni u oksidaciju endogenih supstrata.
Na najvišoj stepenici evolucijske ljestvice je monooksigenazni sustav jetrenih mikrosoma.
P450 igraju važnu ulogu u oksidaciji brojnih spojeva, endogenih (steroidi, žučne kiseline, masne kiseline, prostaglandini, leukotrieni, biogeni amini) i egzogenih (lijekovi, otrovi, proizvodi industrijskog onečišćenja, pesticidi, kancerogeni, mutageni, itd.) itd.), potonji se nazivaju ksenobiotici.
Na temelju tipa kataliziranih reakcija, P450 se može klasificirati kao vanjski tip monooksigenaze. U prisutnosti donora elektrona (NAD(P)H), P450 je sposoban aktivirati molekularni kisik, čiji se jedan atom zatim ugrađuje u molekulu oksidiranog supstrata, a drugi se reducira u vodu."
R + AH + O L ROH + A + H O gdje je R supstrat, ROH je produkt, AH je donor elektrona.
Reakcije oksigenaze katalizirane citokromom P450 vrlo su raznolike. Jedna od najraširenijih reakcija oksidacije ksenobiotika je reakcija oksidativne dealkilacije, koju prati oksidacija alkilne skupine vezane na N-, O- ili S-atome. Drugo mjesto po zastupljenosti zauzimaju reakcije hidroksilacije cikličkih spojeva, koje uključuju hidroksilaciju aromatskih, zasićenih i heterocikličkih ugljikovodika. P450 također može katalizirati reakcije hidroksilacije alifatskih spojeva, N-oksidaciju, oksidativnu deaminaciju i reakcije redukcije azo i nitro spojeva. Reakcije oksidacije prirodnih spojeva uključuju w-oksidaciju zasićenih masnih kiselina, hidroksilaciju steroidnih hormona, žučnih kiselina i kolesterola, biosintezu prostaglandina, peroksidaciju nezasićenih masnih kiselina.
Za razliku od ostalih hemoproteina, koji obično imaju samo jednu aktivnost i strogo definiranu funkciju u stanici, P450, uz monooksigenazu, može ispoljavati i oksidaznu aktivnost, generirajući reaktivne kisikove spojeve u obliku superoksidnih i hidroksilnih radikala, vodikovog peroksida. U tom smislu, u literaturi se P450 ponekad naziva oksidaza miješane funkcije. A.I. Arčakov i sur. otkrili su da P450 također može funkcionirati kao prava oksidaza s četiri elektrona, stvarajući samo vodu iz molekule kisika. P450 također pokazuje aktivnost peroksidaze, koristeći organske perokside ili vodikov peroksid kao kosupstrate u reakciji oksidacije, umjesto NAD(P)H. Postoje dokazi da P450 može katalizirati reakcije dioksigenaze. Dakle, karakteristična značajka P450 je njegova višestrukost funkcija, ali glavna je monooksigenaza. Na sl. Slika 1.32 prikazuje opći dijagram reakcija oksigenaze i oksidaze. U 1. fazi ciklusa oksigenaze (ciklus a), supstrati se vežu za oksidirani oblik P450 i tvore komplekse enzim-supstrat. U tom slučaju, ovisno o podlozi, mogu se pojaviti tri vrste spektralnih promjena: I, II i modificirani II, karakterizirane u diferencijalnom apsorpcijskom spektru maksimumom i minimumom na određenim valnim duljinama. Supstrati tipa I stupaju u interakciju uglavnom s niskospinskim oblikom P450 i atom željeza prelazi iz šest koordiniranog stanja niskog spina u pet koordiniranih stanja visokog spina. U stvaranju kompleksa tipa I vodeću ulogu imaju hidrofobne interakcije nepolarnih supstrata s aktivnim središtem enzima. Kompleksi tipa II nastaju kao rezultat interakcije amino skupine supstrata s atomom željeza hema, koji je ili u visokospinskom ili niskospinskom stanju. U tom slučaju, visokospinski oblik željeza prelazi u niskospinski oblik. Hemsko željezo u takvim je kompleksima u šesterokoordiniranom stanju, a mjesto vezanja kisika zauzima dušik supstrata. Modificirane spektralne promjene tipa II rezultat su interakcije hidroksilne skupine supstrata s visokospinskim oblikom željeza. Stopa interakcije supstrata tipa I s P450 u pravilu je za red veličine veća od one tipa II. U 2. fazi ciklusa monooksigenaze obnavlja se kompleks P450-supstrat. Elektron za redukciju citokroma P450 dolazi iz NADPH-specifičnog flavoproteina. U sljedećim fazama dolazi do aktivacije kisika. Ove faze karakterizira sekvencijalno stvaranje oksi- i peroksi-kompleksa P450. Oksikompleks P450 sposoban je disocirati s otpuštanjem superoksidnih radikala, iz kojih nastaje vodikov peroksid u reakciji dismutacije (ciklus b). Redukcija oksi kompleksa s drugim elektronom dovodi do stvaranja dvoelektronski reduciranog peroksi kompleksa. Vjeruje se da je ovaj stupanj ograničavajući u ciklusu monooksigenaze. Tijekom razgradnje peroksi kompleksa stvara se vodikov peroksid (ciklus c) i formira se reaktivna oksenoidna čestica (FeO) koja sadrži atom kisika sa šest elektrona ligandiran s feri željezom. Atom kisika iz ove čestice može se prenijeti na C-H vezu supstrata i umetnuti u njega. Drugi predloženi mehanizam je mogućnost acilacije distalnog atoma kisika vezanog na hem željezo. Razgradnja ovog kompleksa dovodi do stvaranja perkiseline na aktivnom mjestu P450. Niska reaktivnost perkiseline zahtijeva dodatnu aktivaciju molekule supstrata. Dvoelektronska redukcija oksenoida dovodi do stvaranja vode iz molekule kisika (ciklus d). Najvjerojatnije ne postoji jedinstveni mehanizam za reakcije katalizirane citokromom P450.
Do danas je identificirano više od 160 različitih gena koji kodiraju P450. Podaci o primarnoj strukturi, specifičnosti supstrata, inducibilnosti, lokalizaciji u stanici, strukturi gena i mnogim drugim svojstvima dani su u računalnoj bazi podataka "Cytochrome P450, Database" (CPD), stvorenoj na Institutu za biomedicinsku kemiju Ruske akademije Medicinske znanosti.
Molek. masa različitih P450 kreće se od 44 do 60 kDa. Hemoproteinski monomeri sastoje se od jednog polipeptidnog lanca koji sadrži od 45 do 55% nepolarnih aminokiselinskih ostataka. U nedostatku deterdženta, citokrom postoji u obliku molekularnih nakupina. težine od 300 do 700 kDa. Potpuni slijed aminokiselina utvrđen je za više od 150 citokroma P450. Najbolje su proučeni CYP2B4 i CYP1A2, izolirani iz mikrosoma jetre kunića nakon indukcije fenobarbitalom, odnosno 3-metilkolantrenom. Molekula CYP2B4 sastoji se od 491 aminokiselinskih ostataka, a CYP1A2 - od 516 aminokiselinskih ostataka. D. Ozols i suradnici 1981. i O. Goto i suradnici 1983. uspoređujući primarnu strukturu hemoproteina obitelji CYP2 i CYP1, otkrili su 2 konzervirana područja