Rakenduslik molekulaarbioloogia. Molekulaarbioloog. Töö kirjeldus Molekulaarbioloogia meetodid
Nukleiinhapete ja valkude biosünteesi uurimise edusammud on viinud paljude suure praktilise tähtsusega meetodite loomiseni meditsiinis, põllumajanduses ja paljudes teistes tööstusharudes.
Pärast geneetilise koodi ning päriliku teabe säilitamise ja rakendamise põhiprintsiipide uurimist jäi molekulaarbioloogia areng soiku, kuna puudusid meetodid, mis võimaldasid geenidega manipuleerida, neid isoleerida ja muuta. Nende meetodite tekkimine toimus 1970.–1980. aastatel. See andis võimsa tõuke selle tänaseni õitseva teadusvaldkonna arengule. Esiteks puudutavad need meetodid üksikute geenide saamist ja nende viimist teiste organismide rakkudesse (molekulaarne kloonimine ja transgenees, PCR), aga ka geenides nukleotiidjärjestuse määramise meetodeid (DNA ja RNA sekveneerimine). Neid meetodeid käsitletakse üksikasjalikumalt allpool. Alustame lihtsaimast põhimeetodist, elektroforeesist, ja liigume seejärel keerukamate meetodite juurde.
DNA ELEKTROFOREES
See on DNA-ga töötamise põhimeetod, mida kasutatakse koos peaaegu kõigi teiste meetoditega soovitud molekulide eraldamiseks ja tulemuste analüüsimiseks. DNA fragmentide eraldamiseks pikkuse järgi kasutatakse geelelektroforeesi. DNA on hape, selle molekulid sisaldavad fosforhappejääke, mis lõhustavad prootoni ja omandavad negatiivse laengu (joonis 1).
Seetõttu sisse elektriväli DNA molekulid liiguvad anoodi – positiivselt laetud elektroodi – poole. See toimub elektrolüüdi lahuses, mis sisaldab laengukandjate ioone, mille tõttu see lahus juhib voolu. Fragmentide eraldamiseks kasutatakse polümeeridest (agaroosist või polüakrüülamiidist) valmistatud tihedat geeli. DNA molekulid "põimuvad" selles, mida rohkem, seda pikemad nad on ja seetõttu liiguvad pikimad molekulid kõige aeglasemalt ja kõige lühemad - kõige kiiremini (joonis 2). Enne või pärast elektroforeesi töödeldakse geeli värvainetega, mis seonduvad DNA-ga ja fluorestseeruvad ultraviolettvalguses, ning saadakse geelis olevate ribade muster (vt joonis 3). Proovis olevate DNA fragmentide pikkuse määramiseks võrreldakse neid markeriga, st standardpikkusega fragmentide komplektiga, mis on paigutatud paralleelselt samale geelile (joonis 4).



Kõige olulisemad vahendid DNA-ga töötamiseks on ensüümid, mis viivad läbi DNA transformatsioone elusrakkudes: DNA polümeraasid, DNA ligaasid ja restriktsiooniendonukleaasid ehk restriktsiooniensüümid. DNA polümeraas Tehakse DNA matriitsi süntees, mis võimaldab DNA-d paljundada katseklaasis. DNA ligaasid DNA molekule kokku õmmelda või neis olevaid lünki parandada. Restriktsiooni endonukleaasid, või piirab, lõigake DNA molekulid rangelt määratletud järjestuste järgi, mis võimaldab teil DNA kogumassist üksikuid fragmente välja lõigata. Need fragmendid võivad mõnel juhul sisaldada üksikuid geene.
piirab
Restriktsiooniensüümide poolt äratuntavad järjestused on sümmeetrilised ja katkestused võivad tekkida sellise järjestuse keskel või nihkega (mõlemas DNA ahelas samas kohas). Tegevusskeem erinevad tüübid restriktaas on näidatud joonisel fig. 1. Esimesel juhul saadakse nn "nürid" otsad ja teisel - "kleepuvad" otsad. Põhja "kleepuvate" otste korral on kett teisest lühem, moodustatakse üheahelaline lõik sümmeetrilise järjestusega, mis on moodustatud mõlemas otsas.
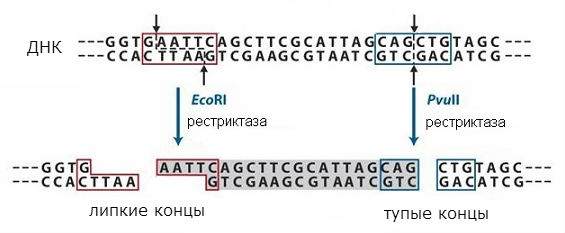
Lõppjärjestused on samad, kui mis tahes DNA lõhustatakse antud restriktsiooniensüümiga ja neid saab uuesti ühendada, kuna neil on komplementaarsed järjestused. Neid saab ligeerida DNA ligaasiga, et moodustada üks molekul. Seega on võimalik kombineerida kahe erineva DNA fragmente ja saada nn rekombinantne DNA. Seda lähenemist kasutatakse molekulaarse kloonimise meetodis, mis võimaldab saada üksikuid geene ja viia need rakkudesse, mis võivad moodustada geenis kodeeritud valku.
molekulaarne kloonimine
Molekulaarsel kloonimisel kasutatakse kahte DNA molekuli – huvipakkuvat geeni sisaldavat inserti ja vektor- DNA toimib kandjana. Insert "õmmeldakse" ensüümide abil vektorisse, saades uue, rekombinantse DNA molekuli, seejärel viiakse see molekul peremeesrakkudesse ja need rakud moodustavad toitekeskkonnal kolooniaid. Koloonia on ühe raku järglane, s.o kloon, kõik koloonia rakud on geneetiliselt identsed ja sisaldavad sama rekombinantset DNA-d. Sellest ka mõiste "molekulaarne kloonimine", st meile huvipakkuvat DNA fragmenti sisaldava rakkude klooni saamine. Kui meid huvitavat sisestust sisaldavad kolooniad on hankitud, saame erinevaid meetodeid selle insertsiooni iseloomustamiseks, näiteks selle täpse järjestuse määramiseks. Rakud võivad toota ka inserdi poolt kodeeritud valku, kui see sisaldab funktsionaalset geeni.
Kui rakkudesse sisestatakse rekombinantne molekul, toimub nende rakkude geneetiline transformatsioon. Muutumine- organismi raku poolt keskkonnast vaba DNA molekuli imendumise protsess ja selle integreerimine genoomi, mis toob kaasa uute pärilike tunnuste ilmnemise sellises rakus, mis on iseloomulikud DNA doonori organismile. . Näiteks kui sisestatud molekul sisaldab antibiootikumi ampitsilliini suhtes resistentsuse geeni, kasvavad transformeeritud bakterid selle juuresolekul. Enne transformatsiooni põhjustas ampitsilliin nende surma, see tähendab, et transformeeritud rakkudesse ilmub uus märk.
VEKTORID
Vektoril peab olema mitmeid omadusi:
Esiteks on see suhteliselt väike DNA molekul, mida on lihtne manipuleerida.
Teiseks, selleks, et DNA rakus säiliks ja paljuneks, peab see sisaldama kindlat järjestust, mis tagab selle replikatsiooni (replikatsiooni alguspunkt ehk replikatsiooni alguspunkt).
Kolmandaks peab see sisaldama marker geen, mis tagab ainult nende rakkude valiku, millesse vektor on sisenenud. Tavaliselt on need antibiootikumiresistentsuse geenid – siis antibiootikumi juuresolekul surevad kõik rakud, mis vektorit ei sisalda.
Geenide kloonimine toimub kõige sagedamini bakterirakkudes, kuna neid on lihtne kultiveerida ja paljuneda kiiresti. Bakterirakus on tavaliselt üks suur ringikujuline, mitme miljoni aluspaari pikkune DNA molekul, mis sisaldab kõiki bakteritele vajalikke geene – bakterikromosoom. Lisaks sellele on mõnes bakteris väike (mitu tuhat aluspaari) ringikujuline DNA, nn plasmiidid(Joonis 2). Need, nagu peamine DNA, sisaldavad nukleotiidjärjestust, mis tagab DNA replikatsioonivõime (ori). Plasmiidid replitseerivad peamisest (kromosomaalsest) DNA-st sõltumatult, seetõttu esinevad nad rakus suure hulga koopiatena. Paljud neist plasmiididest kannavad antibiootikumiresistentsuse geene, mis võimaldab eristada plasmiidi kandvaid rakke normaalsetest rakkudest. Sagedamini kasutatakse plasmiide, mis kannavad kahte geeni, mis annavad resistentsuse kahe antibiootikumi, nagu tetratsükliin ja amütsiliin, suhtes. Sellise plasmiidse DNA eraldamiseks bakteri peamise kromosoomi DNA-st on olemas lihtsad meetodid.

TRANSGENEESI TÄHENDUS
Geenide ülekandumist ühest organismist teise nimetatakse transgenees, ja nii modifitseeritud organismid - transgeensed. Geeniülekande meetodit mikroobirakkudesse kasutatakse meditsiinis kasutatavate rekombinantsete valgupreparaatide saamiseks, eriti inimese valkude, mis ei põhjusta immuunsüsteemi äratõukereaktsiooni - interferoonid, insuliin ja muud valguhormoonid, raku kasvufaktorid, aga ka valkude saamiseks. vaktsiinid. Keerulisematel juhtudel, kui valkude modifitseerimine toimub õigesti ainult eukarüootsetes rakkudes, kasutatakse transgeenseid rakukultuure või transgeenseid loomi, eelkõige kariloomi (peamiselt kitsesid), kes eritavad piima vajalikke valke või eraldatakse nende verest valgud. . Nii saadakse antikehad, vere hüübimisfaktorid ja muud valgud. Transgeneesi meetodil saadakse kultuurtaimed, mis on resistentsed herbitsiidide ja kahjurite suhtes ning millel on muid kasulikud omadused. Kasutades transgeenseid mikroorganisme reovee puhastamiseks ja reostuse vastu võitlemiseks, leidub isegi transgeenseid mikroobe, mis võivad õli lagundada. Lisaks on transgeensed tehnoloogiad teadusuuringutes asendamatud – bioloogia areng pole tänapäeval mõeldav ilma geenide modifitseerimise ja ülekande meetodite rutiinse kasutamiseta.
molekulaarse kloonimise tehnoloogia
lisad
Individuaalse geeni saamiseks mis tahes organismist eraldatakse sellest kogu kromosomaalne DNA ja lõigatakse see ühe või kahe restriktsiooniensüümiga. Ensüümid valitakse nii, et need ei lõikaks ära meile huvipakkuvat geeni, vaid teeksid selle servadest katkestusi ja plasmiidses DNA-s teeksid ühe katkestuse ühes resistentsusgeenis, näiteks ampitsilliini suhtes.
Molekulaarne kloonimise protsess hõlmab järgmisi samme:
Lõika ja õmble – ühe rekombinantse molekuli konstrueerimine inserdist ja vektorist.
Transformatsioon on rekombinantse molekuli viimine rakkudesse.
Valik - rakkude valimine, mis said vektori insertiga.
lõikamine ja õmblemine
Plasmiidi DNA-d töödeldakse samade restriktsiooniensüümidega ja see muutub lineaarseks molekuliks, kui valitakse selline restriktsiooniensüüm, mis viib plasmiidi sisse 1 katkestuse. Selle tulemusena ilmuvad kõigi saadud DNA fragmentide otstesse samad kleepuvad otsad. Temperatuuri langetamisel liituvad need otsad juhuslikult ja ligeeritakse DNA ligaasiga (vt joonis 3).

Saadakse erineva koostisega tsirkulaarsete DNA-de segu: mõned neist sisaldavad teatud kromosomaalse DNA DNA järjestust, mis on seotud bakteri DNA-ga, teised sisaldavad omavahel ühendatud kromosomaalse DNA fragmente ja kolmandad sisaldavad redutseeritud ringikujulist plasmiidi või selle dimeeri. (joonis 4).

muutumine
Järgmisena viiakse see segu läbi geneetiline transformatsioon bakterid, mis ei sisalda plasmiide. Muutumine- organismi raku poolt keskkonnast vaba DNA molekuli imendumise protsess ja selle integreerimine genoomi, mis toob kaasa uute pärilike tunnuste ilmnemise sellises rakus, mis on iseloomulikud DNA doonori organismile. . Igasse rakku saab siseneda ja paljuneda ainult üks plasmiid. Sellised rakud asetatakse tahkele toitekeskkonnale, mis sisaldab antibiootikumi tetratsükliini. Rakud, kes plasmiidi ei saanud, sellel söötmel ei kasva ning plasmiidi kandvad rakud moodustavad kolooniaid, millest igaüks sisaldab vaid ühe raku järglasi, s.o. kõik koloonia rakud kannavad sama plasmiidi (vt joonis 5).

Valik
Järgmiseks on ülesandeks isoleerida ainult need rakud, millesse on sisestatud vektor sisenenud, ja eristada neid rakkudest, mis kannavad ainult vektorit ilma insertita või ei kanna vektorit üldse. Seda õigete lahtrite valimise protsessi nimetatakse valik. Selleks taotlege selektiivsed markerid- tavaliselt antibiootikumiresistentsuse geenid vektoris ja selektiivne meedia mis sisaldavad antibiootikume või muid selektiivseid aineid.
Vaadeldavas näites kultiveeritakse ampitsilliini juuresolekul kasvatatud kolooniate rakke kahel söötmel: esimene sisaldab ampitsilliini ja teine tetratsükliini. Ainult plasmiidi sisaldavad kolooniad kasvavad mõlemal söötmel, samas kui kolooniad, mis sisaldavad plasmiididesse sisestatud kromosomaalset DNA-d, ei kasva tetratsükliiniga söötmel (joonis 5). Nende hulgast valitakse spetsiaalsete meetoditega välja need, mis sisaldavad meile huvipakkuvat geeni, kasvatatakse piisavas koguses ja eraldatakse plasmiidne DNA. Sellest lõigatakse huvipakkuv individuaalne geen välja, kasutades samu restriktaase, mida kasutati rekombinantse DNA saamiseks. Selle geeni DNA-d saab kasutada nukleotiidide järjestuse määramiseks, uute omaduste saamiseks mis tahes organismi sisestamiseks või soovitud valgu sünteesimiseks. Seda geeniisolatsiooni meetodit nimetatakse molekulaarne kloonimine.
fluorestseeruvad valgud
Eukarüootsete organismide uuringutes on väga mugav kasutada fluorestseeruvaid valke markergeenidena. Esimese fluorestseeruva valgu geen, roheline fluorestsentsvalk (GFP) eraldati meduusist Aqeuorea victoria ja viidi erinevatesse mudelorganismidesse (vt joonis 6) 2008. aastal said O. Shimomura, M. Chalfi ja R. Tsien selle valgu avastamise ja rakendamise eest Nobeli preemia.

Seejärel eraldati teiste fluorestseeruvate valkude – punase, sinise, kollase – geenid. Neid geene on kunstlikult modifitseeritud, et toota valke koos soovitud omadused. Fluorestseeruvate valkude mitmekesisus on näidatud joonisel fig. 7, mis näitab Petri tassi bakteritega, mis sisaldavad erinevate fluorestseeruvate valkude geene.

fluorestseeruvate valkude rakendamine
Fluorestseeruva valgu geeni saab liita mis tahes muu valgu geeniga, siis tekib translatsiooni käigus üks valk – translatsiooniline liitvalk või sulandumine(liitvalk), mis fluorestseerib. Seega on võimalik uurida näiteks mis tahes huvipakkuvate valkude paiknemist (asukohta) rakus, nende liikumist. Kasutades fluorestseeruvate valkude ekspressiooni ainult teatud tüüpi rakkudes, on võimalik seda tüüpi rakke markeerida mitmerakulises organismis (vt joonis 8 - hiire aju, milles üksikutel neuronitel on erinevad värvid fluorestseeruvate valgugeenide teatud kombinatsiooni tõttu). Fluorestseeruvad valgud on kaasaegses molekulaarbioloogias asendamatud vahendid.

PCR
Teist meetodit geenide saamiseks nimetatakse polümeraasi ahelreaktsioon (PCR). See põhineb DNA polümeraaside võimel täiendada DNA teist ahelat mööda komplementaarset ahelat, nagu see toimub rakkudes DNA replikatsiooni ajal.
Selle meetodi replikatsiooni alguspunktid annavad kaks väikest DNA tükki, mida nimetatakse seemned, või praimerid. Need praimerid on komplementaarsed kahe DNA ahela huvipakkuva geeni otstega. Esiteks segatakse kromosomaalne DNA, millest geen eraldatakse, seemnetega ja kuumutatakse temperatuurini 99 °C. See toob kaasa vesiniksidemete katkemise ja DNA ahelate lahknemise. Pärast seda alandatakse temperatuuri umbes 50-70 C-ni (olenevalt seemnete pikkusest ja järjestusest). Nendel tingimustel kinnituvad praimerid kromosomaalse DNA komplementaarsete piirkondade külge, moodustades korrapärase topeltheeliksi (vt joonis 9). Pärast seda lisatakse kõigi nelja DNA sünteesiks vajaliku nukleotiidi ja DNA polümeraasi segu. Ensüüm pikendab praimereid, ehitades kaheahelalise DNA praimerite kinnituskohast, st. geeni otstest kuni üheahelalise kromosoomimolekuli lõpuni. 
Kui segu nüüd uuesti kuumutada, hajuvad kromosomaalsed ja äsja sünteesitud ahelad. Pärast jahutamist liituvad nendega uuesti seemned, mida võetakse suures koguses (vt joonis 10).

Äsja sünteesitud ahelatel ühinevad nad mitte selle otsaga, millest esimene süntees algas, vaid vastupidisega, kuna DNA ahelad on antiparalleelsed. Seetõttu valmib sellistel ahelatel teises sünteesitsüklis ainult geenile vastav järjestus (vt joonis 11).

Selle meetodi puhul kasutatakse termofiilsete bakterite DNA polümeraasi, mis talub keemist ja töötab temperatuuril 70–80 °C, seda pole vaja iga kord lisada, vaid piisab, kui lisada katse alguses. Korrates kuumutamise ja jahutamise protseduure samas järjestuses, saame kahekordistada järjestuste arvu igas tsüklis, mis on mõlemast otsast piiratud sisestatud seemnetega (vt joonis 12).

Umbes 25 sellise tsükli järel suureneb geeni koopiate arv enam kui miljon korda. Selliseid koguseid saab katseklaasi sisestatud kromosomaalsest DNA-st kergesti eraldada ja kasutada erinevatel eesmärkidel.
DNA sekveneerimine
Teine oluline saavutus on DNA nukleotiidide järjestuse määramise meetodite väljatöötamine - DNA sekveneerimine(inglise keelest järjestus - järjestus). Selleks on vaja saada muust DNA-st puhtad geenid, kasutades ühte kirjeldatud meetoditest. Seejärel eraldatakse DNA ahelad kuumutamise teel ja neile lisatakse radioaktiivse fosforiga märgistatud praimer või fluorestseeruv märgis. Pange tähele, et võetakse üks seeme, mis täiendab ühte ketti. Seejärel lisatakse DNA polümeraas ja 4 nukleotiidi segu. Selline segu jagatakse 4 osaks ja igaühele lisatakse üks nukleotiididest, modifitseeritud nii, et see ei sisaldaks hüdroksüülrühma kolmandal desoksüriboosi aatomil. Kui selline nukleotiid sisaldub sünteesitud DNA ahelas, siis selle pikenemine ei saa jätkuda, sest polümeraasil pole kuhugi järgmist nukleotiidi kinnitada. Seetõttu katkeb DNA süntees pärast sellise nukleotiidi lisamist. Neid nukleotiide, mida nimetatakse dideoksünukleotiidideks, lisatakse tavapärasest palju vähem, mistõttu ahela lõpetamine toimub ainult aeg-ajalt ja igas ahelas erinevates kohtades. Tulemuseks on ahelate segu erinevad pikkused, igaühe lõpus on sama nukleotiid. Seega vastab ahela pikkus uuritava järjestuse nukleotiidide arvule, näiteks kui meil oli adenüüldideoksünukleotiid ja saadud ahelad olid 2, 7 ja 12 nukleotiidi pikkused, siis adeniin oli 2., seitsmendal ja kaheteistkümnendal positsioonil. geen. Saadud ahelate segu saab elektroforeesi abil hõlpsasti suuruse järgi eraldada ja sünteesitud ahelaid saab radioaktiivsuse järgi tuvastada röntgenfilmil (vt joonis 10).

Selgub pildi allservas näidatud pilt, mida nimetatakse radioautogrammiks. Liikudes mööda seda alt üles ja lugedes iga tsooni veergude kohal olevat tähte, saame autogrammist paremal oleval joonisel näidatud nukleotiidjärjestuse. Selgus, et sünteesi ei peata mitte ainult dideoksünukleotiidid, vaid ka nukleotiidid, milles suhkru kolmandale positsioonile on kinnitunud mingi keemiline rühm, näiteks fluorestseeruv värvaine. Kui iga nukleotiid on märgistatud oma värviga, siis sünteesitud ahelate eraldamisel saadud tsoonid helendavad erineva valgusega. See võimaldab reaktsiooni ühes katseklaasis üheaegselt läbi viia kõigi nukleotiidide jaoks ja eraldades saadud ahelad pikkuse järgi, identifitseerida nukleotiidid värvi järgi (vt joonis 11).

Sellised meetodid võimaldasid määrata mitte ainult üksikute geenide järjestusi, vaid lugeda ka terveid genoome. Nüüd on välja töötatud veelgi kiiremad meetodid geenide nukleotiidjärjestuste määramiseks. Kui esimese inimgenoomi dešifreeris suur rahvusvaheline konsortsium esimesel etteantud meetodil 12 aastaga, teise, teist kasutades kolme aastaga, siis nüüd saab seda teha kuu ajaga. See võimaldab ennustada inimese eelsoodumust paljudele haigustele ja võtta eelnevalt meetmeid nende vältimiseks.
Koomiksiraamat konkursile "bio/mol/text": Täna juhatab molekulaarbioloog Katseklaas teid läbi hämmastava teaduse – molekulaarbioloogia – maailma! Alustame ajaloolise ekskursiooniga läbi selle arenguetappide, kirjeldame peamisi avastusi ja katsetusi alates 1933. aastast. Ja kirjeldame selgelt ka molekulaarbioloogia peamisi meetodeid, mis võimaldasid geenidega manipuleerida, neid muuta ja isoleerida. Nende meetodite ilmumine andis tugeva tõuke molekulaarbioloogia arengule. Ja meenutagem ka biotehnoloogia rolli ja puudutagem üht selle valdkonna populaarseimat teemat – genoomi redigeerimist CRISPR/Cas süsteemide abil.
Konkursi peasponsor ja Skoltechi nominatsiooni partner on .
Konkursi sponsor on firma Diaem: suurim bioloogiliste uuringute ja tootmise seadmete, reaktiivide ja kulumaterjalide tarnija.

Ettevõte sponsoreeris publiku valiku auhinda.
Konkursi "Raamat" sponsor - "Alpina non-fiction"
1. Sissejuhatus. Molekulaarbioloogia olemus

See uurib organismide elutegevuse põhitõdesid makromolekulide tasemel. Molekulaarbioloogia eesmärk on teha kindlaks nende makromolekulide roll ja toimimismehhanismid nende struktuuride ja omadustega seotud teadmiste põhjal.
Ajalooliselt tekkis molekulaarbioloogia nukleiinhappeid ja valke uurivate biokeemia valdkondade väljatöötamise käigus. Kuigi biokeemia uurib ainevahetust, keemiline koostis elusrakud, organismid ja neis läbiviidavad keemilised protsessid, molekulaarbioloogia keskendub geneetilise informatsiooni edasikandumise, paljunemise ja säilitamise mehhanismide uurimisele.
Ja molekulaarbioloogia uurimisobjektiks on nukleiinhapped ise - desoksüribonukleiinhape (DNA), ribonukleiinhape (RNA) - ja valgud, aga ka nende makromolekulaarsed kompleksid - kromosoomid, ribosoomid, multiensüümsüsteemid, mis tagavad valkude ja nukleiinhapete biosünteesi. Molekulaarbioloogia piirneb ka uurimisobjektidega ning ühtib osaliselt molekulaargeneetika, viroloogia, biokeemia ja mitmete teiste seotud bioloogiateadustega.
2. Ajalooline ekskursioon läbi molekulaarbioloogia arenguetappide

Eraldi biokeemia valdkonnana hakkas molekulaarbioloogia arenema eelmise sajandi 30ndatel. Juba siis tekkis vajadus mõista elu fenomeni molekulaarsel tasandil, et uurida geneetilise informatsiooni edastamise ja säilitamise protsesse. Just sel ajal pandi paika molekulaarbioloogia ülesanne valkude ja nukleiinhapete omaduste, struktuuri ja vastastikmõju uurimisel.
Mõistet "molekulaarbioloogia" kasutati esmakordselt aastal 1933 aasta William Astbury fibrillaarsete valkude (kollageen, vere fibriin, kontraktiilsed lihasvalgud) uurimisel. Astbury uuris seost nende valkude molekulaarstruktuuri ja bioloogiliste, füüsikaliste omaduste vahel. Molekulaarbioloogia tekkimise alguses peeti RNA-d ainult taimede ja seente komponendiks ning DNA-d ainult loomadeks. Ja sisse 1935 Andrei Belozersky poolt herneste DNA avastamine tõi kaasa fakti, et DNA sisaldub igas elusrakus.
IN 1940 Kolossaalne saavutus oli George Beadle'i ja Edward Tathami geenide ja valkude vahelise põhjusliku seose tuvastamine. Teadlaste hüpotees "Üks geen – üks ensüüm" pani aluse arusaamale, et valgu spetsiifilist struktuuri reguleerivad geenid. Arvatakse, et geneetilist teavet kodeerib DNA-s spetsiaalne nukleotiidide järjestus, mis reguleerib valkude esmast struktuuri. Hiljem tõestati, et paljudel valkudel on kvaternaarne struktuur. Selliste struktuuride moodustamisel osalevad mitmesugused peptiidahelad. Sellest lähtuvalt on geeni ja ensüümi vahelise seose sätet mõnevõrra muudetud ning nüüd kõlab see nagu "Üks geen – üks polüpeptiid".
IN 1944 1999. aastal tõestasid Ameerika bioloog Oswald Avery ja tema kolleegid (Colin McLeod ja McLean McCarthy), et aine, mis põhjustab bakterite transformatsiooni, on DNA, mitte valgud. Eksperiment tõestas DNA rolli päriliku teabe edastamisel, jättes välja aegunud teadmised geenide valgulise olemuse kohta.

1950. aastate alguses näitas Frederick Sanger, et valguahel on ainulaadne aminohappejääkide järjestus. IN 1951 Ja 1952 aastatel määras teadlane kahe polüpeptiidahela - veise insuliini - täieliku järjestuse IN(30 aminohappejääki) ja A(vastavalt 21 aminohappejääki).
Umbes samal ajal, sisse 1951–1953 Erwin Chargaff sõnastas DNA lämmastikualuste suhte reeglid. Reegli järgi on sõltumata elusorganismide liigierinevusest nende DNA-s adeniini (A) kogus võrdne tümiini (T) ja guaniini (G) kogus tsütosiini kogusega. (C).
IN 1953 tõestas DNA geneetilist rolli. James Watson ja Francis Crick tegid Rosalind Franklini ja Maurice Wilkinsi DNA röntgenpildi põhjal kindlaks DNA ruumilise struktuuri ja esitasid hiljem kinnitatud oletuse selle replikatsiooni (kahekordistumise) mehhanismi kohta, mis on pärilikkuse aluseks.

1958 aasta - molekulaarbioloogia keskse dogma kujunemine Francis Cricki poolt: geneetilise informatsiooni ülekanne läheb DNA → RNA → valgu suunas.
Dogma olemus seisneb selles, et rakkudes toimub teatav suunatud infovoog DNA-st, mis omakorda on algne geneetiline tekst, mis koosneb neljast tähest: A, T, G ja C. See on kirjutatud DNA-sse. topeltheeliks nende tähtede järjestuste kujul - nukleotiidid.
Seda teksti transkribeeritakse. Ja protsessi nimetatakse transkriptsioon. Selle protsessi käigus sünteesitakse RNA, mis on identne geneetilise tekstiga, kuid erinevusega: RNA-s on T asemel U (uratsiil).
Seda RNA-d nimetatakse sõnumitooja RNA (mRNA), või maatriks (mRNA). Saade mRNA viiakse läbi geneetilise koodi abil nukleotiidide triplettjärjestuse kujul. Selle protsessi käigus tõlgitakse DNA ja RNA nukleiinhapete tekst neljatähelisest tekstist kahekümnetäheliseks aminohapete tekstiks.
Looduslikke aminohappeid on vaid kakskümmend ja nukleiinhapete tekstis on neli tähte. Seetõttu toimub geneetilise koodi kaudu tõlge neljatähelisest tähestikust kahekümnetähelisele tähestikule, milles igale kolmele nukleotiidile vastab aminohape. Nii et neljast tähest saab teha tervelt 64 kolmetähelist kombinatsiooni, pealegi on aminohappeid 20. Siit järeldub, et geneetilisel koodil peab tingimata olema degeneratsiooni omadus. Ent tollal polnud geneetilist koodi veel teada, pealegi polnud seda hakatudki dešifreerima, vaid Crick oli oma keskse dogma juba sõnastanud.
Sellegipoolest oli kindel, et kood peab olemas olema. Selleks ajaks oli tõestatud, et sellel koodil oli kolmikmärk. See tähendab, et nukleiinhapetes on konkreetselt kolm tähte ( koodonid) vastavad mis tahes aminohappele. Neid koodoneid on 64, need kodeerivad 20 aminohapet. See tähendab, et iga aminohape vastab korraga mitmele koodonile.
Seega võime järeldada, et keskne dogma on postulaat, mis ütleb, et rakus toimub suunatud infovoog: DNA → RNA → valk. Crick rõhutas keskse dogma põhisisu: vastupidist infovoogu ei saa toimuda, valk ei ole võimeline geneetilist informatsiooni muutma.
See on keskse dogma peamine tähendus: valk ei ole võimeline muutma ja transformeerima teavet DNA-ks (või RNA-ks), voog läheb alati ainult ühes suunas.
Mõni aeg pärast seda avastati uus ensüüm, mida keskse dogma koostamise ajal ei tuntud, - pöördtranskriptaas mis sünteesib RNA-st DNA-d. Ensüüm avastati viirustes, mille geneetiline informatsioon on kodeeritud RNA-s, mitte DNA-s. Selliseid viiruseid nimetatakse retroviirusteks. Neil on viiruskapsel, millesse on suletud RNA ja spetsiaalne ensüüm. Ensüüm on pöördtranskriptaas, mis sünteesib DNA-d vastavalt selle viiruse RNA mallile ja see DNA toimib seejärel geneetilise materjalina viiruse edasiseks arendamiseks rakus.
Muidugi põhjustas see avastus molekulaarbioloogide seas suure šoki ja palju poleemikat, kuna usuti, et keskse dogma põhjal ei saa see nii olla. Crick selgitas aga kohe, et ta pole kunagi öelnud, et see on võimatu. Ta ütles vaid, et kunagi ei saa toimuda infovoogu valgult nukleiinhapetele ja juba nukleiinhapete sees on igasugused protsessid täiesti võimalikud: DNA süntees DNA-l, DNA süntees RNA-l, RNA süntees DNA-l ja RNA süntees RNA-l.
Pärast keskse dogma sõnastamist jäi õhku veel hulk küsimusi: kuidas kodeerib DNA (või RNA) moodustava nelja nukleotiidi tähestik valke moodustavate aminohapete 20-tähelist tähestikku? Mis on geneetilise koodi olemus?
Esimesed ideed geneetilise koodi olemasolu kohta sõnastas Alexander Downes ( 1952 d.) ja Georgi Gamov ( 1954 G.). Teadlased on näidanud, et nukleotiidide järjestus peab sisaldama vähemalt kolme linki. Hiljem tõestati, et selline järjestus koosneb kolmest nukleotiidist, nn koodon (kolmik). Kuid küsimus, millised nukleotiidid vastutavad millise aminohappe valgu molekuli lisamise eest, jäi lahtiseks kuni 1961. aastani.

Ja sisse 1961 Marshall Nirenberg ja Heinrich Mattei kasutasid seda süsteemi ringhäälingu edastamiseks in vitro. Matriitsina kasutati oligonukleotiidi. See sisaldas ainult uratsiili jääke ja sellest sünteesitud peptiid sisaldas ainult aminohapet fenüülalaniini. Seega tehti esmalt kindlaks koodoni tähendus: koodon UUU kodeerib fenüülalaniini. Hiljem leidis Har Koraan, et nukleotiidjärjestus UCUCUCUCUCUCUC kodeerib aminohapete komplekti seriin-leutsiin-seriin-leutsiin. Üldiselt tänu Nirenbergi ja Koraani teostele 1965 aastal oli geneetiline kood täielikult lahti harutatud. Selgus, et iga kolmik kodeerib kindlat aminohapet. Ja koodonite järjekord määrab valgu aminohapete järjekorra.
Valkude ja nukleiinhapete toimimise põhiprintsiibid sõnastati 70ndate alguseks. Leiti, et valkude ja nukleiinhapete süntees toimub maatriksmehhanismi järgi. Matriitsmolekul kannab kodeeritud teavet aminohapete või nukleotiidide järjestuse kohta. Replikatsiooni või transkriptsiooni ajal on matriitsiks DNA ning translatsiooni ja pöördtranskriptsiooni ajal mRNA.
Nii loodi eeldused molekulaarbioloogia, sh geenitehnoloogia valdkondade kujunemiseks. Ja 1972. aastal töötasid Paul Berg ja kolleegid välja molekulaarse kloonimise tehnoloogia. Teadlased on saanud esimese rekombinantse DNA in vitro. Need silmapaistvad avastused moodustasid aluse molekulaarbioloogias uuele suunale ja 1972 aastat on sellest ajast peale peetud geenitehnoloogia sünnikuupäevaks.



3. Molekulaarbioloogia meetodid

Tohutud edusammud nukleiinhapete, DNA struktuuri ja valkude biosünteesi uurimisel on viinud mitmete meditsiinis, põllumajanduses ja teaduses üldiselt väga oluliste meetodite loomiseni.
Pärast geneetilise koodi ja päriliku teabe säilitamise, edastamise ja rakendamise põhiprintsiipide uurimist muutusid molekulaarbioloogia edasiseks arenguks vajalikuks spetsiaalsed meetodid. Need meetodid võimaldaksid geene manipuleerida, muuta ja isoleerida.
Selliste meetodite tekkimine toimus 1970. ja 1980. aastatel. See andis tohutu tõuke molekulaarbioloogia arengule. Esiteks on need meetodid otseselt seotud geenide tootmise ja nende viimisega teiste organismide rakkudesse, samuti nukleotiidjärjestuse määramise võimalusega geenides.
3.1. DNA elektroforees

DNA elektroforees on DNA-ga töötamise põhimeetod. DNA elektroforeesi kasutatakse koos peaaegu kõigi teiste meetoditega soovitud molekulide eraldamiseks ja tulemuste edasiseks analüüsimiseks. Geelelektroforeesi meetodit kasutatakse DNA fragmentide eraldamiseks pikkuse järgi.


Enne või pärast elektroforeesi töödeldakse geeli värvainetega, mis võivad DNA-ga seonduda. Värvained fluorestseerivad ultraviolettvalguses, mille tulemuseks on geelis ribade muster. DNA fragmentide pikkuse määramiseks saab neid võrrelda markerid- standardpikkusega fragmentide komplektid, mis kantakse samale geelile.

Fluorestseeruvad valgud
Eukarüootsete organismide uurimisel on mugav kasutada markergeenidena fluorestseeruvaid valke. Esimese rohelise fluorestseeruva valgu geen ( roheline fluorestseeruv valk, GFP) isoleeritud millimallikast Aqeuorea victoria ja seejärel viidud erinevatesse organismidesse. Pärast seda eraldati teiste värvide fluorestseeruvate valkude geenid: sinine, kollane, punane. Huvipakkuvate omadustega valkude saamiseks on selliseid geene kunstlikult modifitseeritud.

Üldiselt on DNA molekuliga töötamiseks kõige olulisemad vahendid ensüümid, mis viivad rakkudes läbi mitmeid DNA transformatsioone: DNA polümeraas, DNA ligaasid Ja piirab (restriktsiooni endonukleaasid).
transgenees
transgenees Seda nimetatakse geenide ülekandmiseks ühelt organismilt teisele. Selliseid organisme nimetatakse transgeensed.
Rekombinantsed valgupreparaadid saadakse lihtsalt geenide ülekandmisel mikroorganismide rakkudesse. Enamik neist valkudest on interferoonid, insuliini, mõned valguhormoonid, aga ka valgud mitmete vaktsiinide tootmiseks.
Muudel juhtudel kasutatakse eukarüootide või transgeensete loomade, enamasti kariloomade rakukultuure, mis eritavad piima vajalikke valke. Nii saadakse antikehad, vere hüübimisfaktorid ja muud valgud. Kahjurite ja herbitsiidide suhtes resistentsete põllukultuuride saamiseks kasutatakse transgeneesi meetodit ning reovee puhastamine toimub transgeensete mikroorganismide abil.
Lisaks kõigele eelnevale on transgeensed tehnoloogiad teadusuuringutes asendamatud, sest bioloogia areng on geenide modifitseerimise ja ülekande meetodite kasutamisega kiirem.

Piiravad
Restriktsiooniensüümide poolt äratuntavad järjestused on sümmeetrilised, seega võivad tekkida igasugused katkestused kas sellise järjestuse keskel või nihkega DNA molekuli ühes või mõlemas ahelas.
Mis tahes DNA tükeldamisel restriktsiooniensüümiga on fragmentide otstes olevad järjestused samad. Nad saavad uuesti ühenduse luua, kuna neil on üksteist täiendavad saidid.
Nende järjestuste õmblemisel saate ühe molekuli DNA ligaasid. Tänu sellele on võimalik kombineerida kahe erineva DNA fragmente ja saada rekombinantne DNA.


3.2. PCR
Meetod põhineb DNA polümeraaside võimel täiendada DNA teist ahelat mööda komplementaarset ahelat samamoodi nagu DNA replikatsiooni protsessis rakus.



3.3. DNA sekveneerimine
Sekveneerimismeetodi kiire areng võimaldab tõhusalt määrata uuritava organismi omadusi selle genoomi tasemel. Selliste genoomiliste ja postgenoomiliste tehnoloogiate peamine eelis on uurimis- ja õppimisvõimaluste suurenemine. geneetiline olemus inimeste haigusi, et ette võtta vajalikke meetmeid ja vältida haigusi.
Suuremahuliste uuringute abil on võimalik saada vajalikke andmeid erinevate inimrühmade erinevate geneetiliste omaduste kohta, arendades seeläbi meditsiini meetodeid. Seetõttu on tänapäeval väga populaarne erinevate haiguste geneetilise eelsoodumuse tuvastamine.
Sarnased meetodid on laialdaselt rakendatavad praktiliselt kogu maailmas, sealhulgas Venemaal. Tänu teaduse arengule võetakse sellised meetodid kasutusele meditsiiniuuringutes ja meditsiinipraktikaüldiselt.



4. Biotehnoloogia

Biotehnoloogia- distsipliin, mis uurib elusorganismide või nende süsteemide kasutamise võimalusi tehnoloogiliste probleemide lahendamiseks, samuti soovitud omadustega elusorganismide loomist geenitehnoloogia abil. Biotehnoloogias rakendatakse keemia, mikrobioloogia, biokeemia ja loomulikult molekulaarbioloogia meetodeid.
Biotehnoloogia arengu põhisuunad (biotehnoloogiliste protsesside põhimõtted juurutatakse kõigis tööstusharudes):
- Uut tüüpi toidu ja loomasööda loomine ja tootmine.
- Uute mikroorganismide tüvede hankimine ja uurimine.
- Uute taimesortide aretamine, samuti vahendite loomine taimede kaitsmiseks haiguste ja kahjurite eest.
- Biotehnoloogia meetodite rakendamine ökoloogia vajadusteks. Selliseid biotehnoloogia meetodeid kasutatakse jäätmete kõrvaldamiseks, puhastamiseks Reovesi, väljatõmbeõhk ja pinnase kanalisatsioon.
- Vitamiinide, hormoonide, ensüümide, seerumite tootmine meditsiini vajadusteks. Biotehnoloogid arenevad paremaks ravimid varem ravimatuks peetud.
Biotehnoloogia suur saavutus on geenitehnoloogia.
Geenitehnoloogia- tehnoloogiate ja meetodite kogum rekombinantsete RNA ja DNA molekulide saamiseks, üksikute geenide eraldamiseks rakkudest, geenidega manipuleerimiseks ja nende viimiseks teistesse organismidesse (bakterid, pärm, imetajad). Sellised organismid on võimelised tootma soovitud muudetud omadustega lõpptooteid.
Geenitehnoloogia meetodid on suunatud uute, varem looduses olematute geenikombinatsioonide konstrueerimisele.
Geenitehnoloogia saavutustest rääkides on võimatu mitte puudutada kloonimise teemat. Kloonimine on üks biotehnoloogia meetoditest, mida kasutatakse mittesugulise paljunemise teel erinevate organismide identsete järglaste saamiseks.
Teisisõnu võib kloonimist käsitleda kui organismi või raku geneetiliselt identsete koopiate loomise protsessi. Ja kloonitud organismid on sarnased või täiesti identsed mitte ainult väliste tunnuste, vaid ka geneetilise sisu poolest.
Kurikuulsast lambast Dollyst sai 1966. aastal esimene kloonitud imetaja. See saadi somaatilise raku tuuma siirdamisel munaraku tsütoplasmasse. Dolly oli tuumadoonorlamba geneetiline koopia. Looduslikes tingimustes moodustub isend ühest viljastatud munarakust, olles saanud poole geneetilisest materjalist kahelt vanemalt. Kloonimise käigus võeti aga geneetiline materjal ühe isendi rakust. Kõigepealt eemaldati sügoodist tuum, mis sisaldab DNA-d ennast. Seejärel eemaldati täiskasvanud lamba rakust tuum ja siirdati see ilma tuumata sigooti, seejärel siirdati see täiskasvanud inimese emakasse ning lasti kasvada ja areneda.

Kõik kloonimiskatsed pole aga olnud edukad. Paralleelselt Dolly kloonimisega viidi läbi DNA asenduskatse 273 teise munaga. Kuid ainult ühel juhul sai täiskasvanud elusloom täielikult areneda ja kasvada. Pärast Dollyt üritasid teadlased kloonida teist tüüpi imetajaid.

Üks geenitehnoloogia liike on genoomi redigeerimine.
CRISPR/Cas tööriist põhineb bakterite immuunkaitsesüsteemi elemendil, mida teadlased on kohandanud loomade või taimede DNA-s mistahes muutuste sisseviimiseks.


CRISPR/Cas on üks biotehnoloogilisi meetodeid üksikute geenide manipuleerimiseks rakkudes. Sellel tehnoloogial on palju rakendusi. CRISPR/Cas võimaldab teadlastel välja selgitada erinevate geenide funktsiooni. Selleks tuleb lihtsalt uuritav geen DNA-st välja lõigata ja uurida, milliseid organismi funktsioone see mõjutas.
Mõned süsteemi praktilised rakendused:
- Põllumajandus. CRISPR/Cas süsteemide abil saab põllukultuure parandada. Just nimelt selleks, et need oleksid maitsvamad ja toitvamad, aga ka kuumakindlad. Taimi on võimalik varustada muude omadustega: näiteks lõigata pähklitest (maapähklitest või sarapuupähklitest) välja allergeenigeen.
- Meditsiin, pärilikud haigused. Teadlaste eesmärk on kasutada CRISPR/Casi, et eemaldada inimese genoomist mutatsioonid, mis võivad põhjustada haigusi, nagu sirprakuline aneemia jne. Teoreetiliselt võib CRISPR/Cas peatada HIV-i arengu.
- Geeniajam. CRISPR/Cas võib muuta mitte ainult üksiku looma või taime genoomi, vaid ka liigi genofondi. Seda mõistet tuntakse kui "geeniajam". Iga elusorganism annab pooled oma geenidest edasi oma järglastele. Kuid CRISPR/Cas kasutamine võib suurendada geeniülekande võimalust kuni 100%. See on oluline soovitud tunnuse kiiremaks levimiseks kogu populatsioonis.
Šveitsi teadlased on oluliselt täiustanud ja moderniseerinud CRISPR/Cas genoomi redigeerimise meetodit, avardades seeläbi selle võimalusi. Kuid teadlased said CRISPR / Cas süsteemi abil korraga muuta ainult ühte geeni. Kuid nüüd on ETH Zürichi teadlased välja töötanud meetodi, mis suudab üheaegselt modifitseerida rakus 25 geeni.
Uusima tehnika jaoks kasutasid eksperdid ensüümi Cas12a. Geneetikud on esimest korda ajaloos edukalt ahve klooninud. "Populaarne mehaanika";
Molekulaarbioloogia on läbi elanud oma uurimismeetodite kiire arengu perioodi, mis nüüd erineb biokeemiast. Nende hulka kuuluvad eelkõige geenitehnoloogia, kloonimise, kunstliku ekspressiooni ja geenide väljalülitamise meetodid. Kuna DNA on geneetilise informatsiooni materiaalne kandja, on molekulaarbioloogia muutunud geneetikale palju lähedasemaks ja ristmikul tekkis molekulaargeneetika, mis on nii geneetika kui ka molekulaarbioloogia osa. Nii nagu molekulaarbioloogia kasutab laialdaselt viiruseid uurimisvahendina, kasutab viroloogia oma probleemide lahendamiseks molekulaarbioloogia meetodeid. Arvutitehnoloogia tegeleb geneetilise informatsiooni analüüsiga, millega seoses on tekkinud uued molekulaargeneetika valdkonnad, mida mõnikord peetakse eridistsipliinideks: bioinformaatika, genoomika ja proteoomika.
Arengu ajalugu
See seemneline avastus valmistati ette viiruste ja bakterite geneetika ja biokeemia pikaajalise uurimistöö käigus.
1928. aastal näitas Frederick Griffith esimest korda, et kuumusega tapetud patogeensete bakterite ekstrakt võib patogeensuse tunnuse üle kanda healoomulistele bakteritele. Bakterite transformatsiooni uurimine viis edasi haigusetekitaja puhastamiseni, mis vastupidiselt ootustele osutus mitte valguks, vaid nukleiinhappeks. Nukleiinhape ise ei ole ohtlik, see kannab ainult geene, mis määravad mikroorganismi patogeensuse ja muud omadused.
XX sajandi 50ndatel näidati, et bakteritel on primitiivne seksuaalprotsess, nad on võimelised vahetama kromosomaalset DNA-d, plasmiide. Plasmiidide avastamine ja ka transformatsioonid olid molekulaarbioloogias levinud plasmiiditehnoloogia aluse. Teine oluline avastus metoodika jaoks oli bakteriaalsete viiruste, bakteriofaagide avastamine 20. sajandi alguses. Faagid võivad kanda ka geneetilist materjali ühest bakterirakust teise. Bakterite nakatumine faagidega põhjustab bakteri RNA koostise muutumise. Kui ilma faagideta on RNA koostis sarnane bakteriaalse DNA koostisega, siis pärast nakatumist muutub RNA sarnasemaks bakteriofaagi DNA-ga. Seega leiti, et RNA struktuuri määrab DNA struktuur. Valkude sünteesi kiirus rakkudes omakorda sõltub RNA-valgu komplekside hulgast. Nii see sõnastati Molekulaarbioloogia keskne dogma: DNA ↔ RNA → valk.
Molekulaarbioloogia edasise arenguga kaasnes nii selle metoodika väljatöötamine, eelkõige DNA nukleotiidjärjestuse määramise meetodi leiutamine (W. Gilbert ja F. Sanger, Nobeli keemiaauhind 1980), kui ka uued. avastused geenide ehituse ja talitluse uurimise vallas (vt. Geneetika ajalugu). 21. sajandi alguseks saadi andmed kogu inimese DNA algstruktuuri ja mitmete teiste organismide, meditsiini jaoks kõige olulisemate organismide kohta, Põllumajandus ja teadusuuringud, mille tulemusel tekkisid bioloogias mitmed uued valdkonnad: genoomika, bioinformaatika jne.
Vaata ka
- Molekulaarbioloogia (ajakiri)
- Transkriptoomika
- Molekulaarpaleontoloogia
- EMBO – Euroopa Molekulaarbioloogia Organisatsioon
Kirjandus
- Laulja M., Berg P. Geenid ja genoomid. - Moskva, 1998.
- Stent G., Kalindar R. Molekulaargeneetika. - Moskva, 1981.
- Sambrook J., Fritsch E.F., Maniatis T. Molekulaarne kloonimine. - 1989.
- Patrušev L.I. Geenide ekspressioon. - M.: Nauka, 2000. - 000 lk., ill. ISBN 5-02-001890-2
Lingid
Wikimedia sihtasutus. 2010 .
- Nižni Novgorodi oblasti Ardatovski rajoon
- Nižni Novgorodi oblasti Arzamasi rajoon
Vaadake, mis on "molekulaarbioloogia" teistes sõnaraamatutes:
MOLEKULAARBIOLOOGIA- õpib põhitõdesid. elu omadused ja ilmingud molekulaarsel tasandil. Olulisemad suunad M. b. on uuringud rakkude geneetilise aparaadi struktuurse ja funktsionaalse korralduse ning päriliku teabe rakendamise mehhanismi kohta ... ... Bioloogia entsüklopeediline sõnastik
MOLEKULAARBIOLOOGIA- uurib elu põhiomadusi ja ilminguid molekulaarsel tasandil. Saab teada, kuidas ja mil määral on organismide kasv ja areng, päriliku teabe talletamine ja edastamine, energia muundumine elusrakkudes ja muud nähtused tingitud ... Suur entsüklopeediline sõnaraamat
MOLEKULAARBIOLOOGIA Kaasaegne entsüklopeedia
MOLEKULAARBIOLOOGIA- MOLEKULAARBIOLOOGIA, elusorganisme moodustavate Molekulide ehituse ja funktsiooni bioloogiline uurimine. Peamised õppesuunad on füüsilised ja Keemilised omadused valgud ja nukleiinhapped nagu DNA. Vaata ka… … Teaduslik ja tehniline entsüklopeediline sõnastik
molekulaarbioloogia- osa biol.-st, mis uurib elu põhiomadusi ja ilminguid molekulaarsel tasandil. Saab teada, kuidas ja mil määral toimub organismide kasv ja areng, päriliku informatsiooni talletamine ja edastamine, energia muundamine elusrakkudes ja ... ... Mikrobioloogia sõnaraamat
molekulaarbioloogia- — Biotehnoloogia teemad ET molekulaarbioloogia … Tehnilise tõlkija käsiraamat
Molekulaarbioloogia- MOLEKULAARBIOLOOGIA, uurib elu põhiomadusi ja ilminguid molekulaarsel tasandil. Saab teada, kuidas ja mil määral toimub organismide kasv ja areng, päriliku informatsiooni talletamine ja edastamine, energia muundamine elusrakkudes ja ... ... Illustreeritud entsüklopeediline sõnaraamat
Molekulaarbioloogia- teadus, mis seab oma ülesandeks elunähtuste olemuse tundmise, uurides bioloogilisi objekte ja süsteeme molekulaarsele tasemele läheneval, mõnel juhul ka selle piirini jõudval tasemel. Selle lõppeesmärk on…… Suur Nõukogude entsüklopeedia
MOLEKULAARBIOLOOGIA- uurib elunähtusi makromolekulide tasemel (ch. arr. valgud ja nukleiinhapped) rakuvabades struktuurides (ribosoomid jne), viirustes, aga ka rakkudes. M. eesmärk. nende makromolekulide rolli ja toimimismehhanismi kindlaksmääramine, mis põhineb ... ... Keemia entsüklopeedia
molekulaarbioloogia- uurib elu põhiomadusi ja ilminguid molekulaarsel tasandil. Saab teada, kuidas ja mil määral toimub organismide kasv ja areng, päriliku informatsiooni talletamine ja edasiandmine, energia muundumine elusrakkudes ja muud nähtused ... ... entsüklopeediline sõnaraamat
Raamatud
- Raku molekulaarbioloogia. Probleemiraamat, J. Wilson, T. Hunt. Ameerika autorite raamat on lisa B. Albertsi, D. Bray, J. Lewise jt õpiku `Molecular Biology of the Cell` 2. väljaandele, mis sisaldab küsimusi ja ülesandeid, mille eesmärk on süvendada . ..
Molekulaarbioloogia, teadus, mis seab oma ülesandeks elunähtuste olemuse tundmise, uurides bioloogilisi objekte ja süsteeme molekulaarsele tasemele läheneval, mõnel juhul ka selle piirini jõudval tasemel. Lõppeesmärk on sel juhul selgitada, kuidas ja mil määral ilmnevad elule iseloomulikud ilmingud, nagu pärilikkus, omasuguste sigimine, valkude biosüntees, erutuvus, kasv ja areng, teabe salvestamine ja edastamine, energia muundumine, liikuvus, jne , on tingitud bioloogiliselt oluliste ainete molekulide struktuurist, omadustest ja vastastikmõjust, peamiselt kahest põhiklassist kõrgmolekulaarsed biopolümeerid – valgud ja nukleiinhapped. Eripäraks M. b. - elutute objektide või elutute objektide elunähtuste uurimine või need, mida iseloomustavad elu kõige primitiivsemad ilmingud. Need on bioloogilised moodustised rakutasandilt ja allapoole: subtsellulaarsed organellid, näiteks isoleeritud rakutuumad, mitokondrid, ribosoomid, kromosoomid, rakumembraanid; edasi - süsteemid, mis seisavad elava ja elutu looduse piiril - viirused, sealhulgas bakteriofaagid ja lõpetades molekulidega kriitilised komponendid elusaine - nukleiinhapped ja valgud.
Vundamendi, millele M. arenes, panid sellised teadused nagu geneetika, biokeemia, elementaarprotsesside füsioloogia jne. Vastavalt oma arengu algallikatele on M. b. on lahutamatult seotud molekulaargeneetikaga, mis on jätkuvalt oluline osa
Eripäraks M. b. on selle kolmemõõtmelisus. M. b. olemus. M. Perutz näeb seda bioloogiliste funktsioonide tõlgendamisel molekulaarstruktuuri kaudu. M. b. teeb oma ülesandeks saada vastused küsimusele "kuidas", teades kogu molekuli struktuuri rolli ja osaluse olemust, ning küsimustele "miks" ja "miks", olles ühelt poolt välja selgitanud , seost molekuli (taas eeskätt valkude ja nukleiinhapete) omaduste ja selle poolt täidetavate funktsioonide vahel ning teisest küljest selliste üksikute funktsioonide rolli elutegevuse ilmingute üldises kompleksis.
Molekulaarbioloogia olulisemad saavutused. Siin on nende saavutuste kaugeltki täielik loetelu: DNA, igat tüüpi RNA ja ribosoomide struktuuri ja bioloogilise funktsiooni mehhanismi avalikustamine, geneetilise koodi avalikustamine; pöördtranskriptsiooni avastamine, st DNA süntees RNA matriitsil; hingamisteede pigmentide toimimismehhanismide uurimine; kolmemõõtmelise struktuuri ja selle funktsionaalse rolli avastamine ensüümide toimes, maatrikssünteesi põhimõte ja valkude biosünteesi mehhanismid; viiruste struktuuri ja nende replikatsioonimehhanismide, antikehade esmase ja osaliselt ruumilise struktuuri avalikustamine; üksikute geenide isoleerimine, keemiline ja seejärel bioloogiline (ensümaatiline) geenide süntees, sh inimese, väljaspool rakku (in vitro); geenide ülekandmine ühest organismist teise, sealhulgas inimese rakkudesse; üha suurema hulga üksikute valkude, peamiselt ensüümide, aga ka nukleiinhapete keemilise struktuuri kiiresti edenev dešifreerimine; mõnede järjest keerukama keerukusega bioloogiliste objektide "isekoosnemise" nähtuste avastamine, alustades nukleiinhappemolekulidest ja liikudes edasi mitmekomponentsete ensüümide, viiruste, ribosoomide jne juurde; allosteeriliste ja muude bioloogiliste funktsioonide ja protsesside reguleerimise aluspõhimõtete selgitamine.
Molekulaarbioloogia probleemid. Koos täpsustatud oluliste ülesannetega oleks M.. ("äratundmise", iseseadumise ja integratsiooni seaduste tundmine) teaduslike otsingute tegelik suund lähitulevikus on struktuuri dešifreerimist võimaldavate meetodite väljatöötamine ja seejärel kõrgmolekulaarsete kolmemõõtmeline, ruumiline organiseerimine. nukleiinhapped. Kõik olulisemad meetodid, mille kasutamine tagas M. b. tekke ja edu, pakkusid välja ja töötasid välja füüsikud (ultratsentrifuugimine, röntgendifraktsioonanalüüs, elektronmikroskoopia, tuumamagnetresonants jne). Peaaegu kõik uued füüsikalised eksperimentaalsed lähenemised (näiteks arvutite, sünkrotroni või bremsstrahlungi, kiirguse, lasertehnoloogia jm kasutamine) avavad uusi võimalusi M. b. probleemide süvauurimiseks. Kõige olulisemate praktilise iseloomuga ülesannete hulgas, millele M. b.-lt vastust oodatakse, on esiteks pahaloomulise kasvu molekulaarse aluse probleem, seejärel - pärilike haiguste ennetamise ja võib-olla nende ületamise viisid - " molekulaarsed haigused". Suur tähtsus on bioloogilise katalüüsi molekulaarse aluse ehk ensüümide toime selgitamisel. Kõige olulisemate hulgas kaasaegsed trendid M. b. peaks sisaldama soovi dešifreerida hormoonide, toksiliste ja raviainete molekulaarseid toimemehhanisme, samuti selgitada välja selliste rakustruktuuride molekulaarstruktuuri ja toimimise üksikasjad nagu bioloogilised membraanid, mis osalevad läbitungimise protsesside reguleerimises ja ainete transport. Kaugemad väravad M. b. - teadmised närviprotsesside olemusest, mälu mehhanismidest jne. Üks olulisi esilekerkivaid sektsioone M. b. - nn. geenitehnoloogia, mis seab oma ülesandeks elusorganismide geneetilise aparaadi (genoomi) sihipärase toimimise, alustades mikroobidest ja madalamatest (üherakulistest) ning lõpetades inimesega (viimasel juhul eelkõige elusorganismide radikaalse ravi eesmärgil). pärilikud haigused ja geneetiliste defektide korrigeerimine).
MB olulisemad suunad:
- Molekulaargeneetika - raku geneetilise aparaadi struktuurse ja funktsionaalse korralduse ning päriliku teabe rakendamise mehhanismi uurimine
– Molekulaarviroloogia – viiruste ja rakkude interaktsiooni molekulaarsete mehhanismide uurimine
– Molekulaarimmunoloogia – keha immuunreaktsioonide mustrite uurimine
- Arengu molekulaarbioloogia - rakkude mitmekesisuse ilmnemise uurimine organismide individuaalse arengu ja rakkude spetsialiseerumise käigus
Peamised uurimisobjektid: Viirused (sh bakteriofaagid), Rakud ja subtsellulaarsed struktuurid, Makromolekulid, Mitmerakulised organismid.