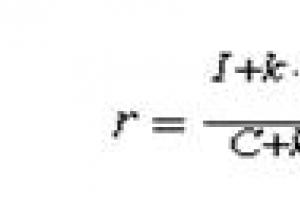rodzaje receptorów. Polimorfia uczuć: jak działają receptory skórne Czym są receptory w definicji biologii
Receptory
Dwa tysiące lat temu Arystoteles napisał, że człowiek ma pięć zmysłów: wzrok, słuch, dotyk, węch i smak. Przez dwa tysiąclecia naukowcy wielokrotnie odkrywali narządy nowych „szóstych zmysłów”, na przykład aparat przedsionkowy czy receptory temperatury. Te narządy zmysłów są często określane jako „bramy do świata”: pozwalają zwierzętom poruszać się w środowisku zewnętrznym i odbierać sygnały od własnego gatunku. Jednak nie mniej ważne w życiu zwierząt jest „zajrzenie w głąb siebie”; naukowcy odkryli różne receptory, które mierzą ciśnienie krwi, poziom cukru we krwi i dwutlenek węgla, ciśnienie osmotyczne krwi, napięcie mięśni itp. Te wewnętrzne receptory, z których sygnały z reguły nie docierają do świadomości, pozwalają naszemu układowi nerwowemu kontrolować szereg procesów zachodzących w organizmie.
Z tego, co zostało powiedziane, jasno wynika, że klasyfikacja Arystotelesa jest wyraźnie przestarzała i dziś liczba różnych „zmysłów” byłaby bardzo duża, zwłaszcza jeśli weźmiemy pod uwagę narządy zmysłów różnych organizmów zamieszkujących Ziemię.
Jednocześnie, badając tę różnorodność, stwierdzono, że praca wszystkich narządów zmysłów opiera się na jednej zasadzie. Wpływ zewnętrzny jest odbierany przez specjalne komórki - receptory i zmienia MP tych komórek. Ten sygnał elektryczny nazywany jest potencjałem receptora. A potem potencjał receptora kontroluje uwalnianie mediatora z komórki receptora, czyli częstotliwość jego impulsów. Tak więc receptor jest konwerterem wpływów zewnętrznych na sygnały elektryczne, jak znakomicie odgadł Volt.
Receptory przekazują sygnały do układu nerwowego, gdzie są dalej przetwarzane.
W dawnych czasach w produkcji przyrządy umieszczano bezpośrednio w punktach pomiarowych. Na przykład każdy kocioł parowy był wyposażony we własny termometr i manometr. Jednak w przyszłości takie urządzenia z reguły zostały zastąpione czujnikami, które przetwarzają temperaturę lub ciśnienie na sygnały elektryczne; sygnały te można było łatwo przesyłać na odległość. Teraz operator patrzy na panel, na którym zmontowane są instrumenty pokazujące temperaturę, ciśnienie, prędkość turbiny itp. i nie powinien obchodzić wszystkich jednostek po kolei. W rzeczywistości żywe organizmy opracowały taki postępowy system pomiaru różnych wielkości setki milionów lat przed nadejściem technologii. Rolę tarczy, która odbiera wszystkie sygnały, pełni mózg.
Naturalne jest klasyfikowanie różnych receptorów według rodzajów postrzeganych przez nie wpływów zewnętrznych. Na przykład tak różne receptory, jak receptory narządu słuchu, receptory narządu równowagi, receptory zapewniające dotyk, reagują na wpływy zewnętrzne tego samego typu - mechaniczne. Z tego punktu widzenia można wyróżnić następujące typy receptorów.
1) Fotoreceptory, komórki reagujące na fale elektromagnetyczne, których częstotliwość mieści się w pewnym zakresie.
2) Mechanoreceptory, komórki reagujące na przemieszczenie ich części względem siebie; mechanoreceptory, jak już wspomniano, obejmują komórki, które odbierają dźwięki, tj. wibracje wody i powietrza o określonej częstotliwości, oraz mechanoreceptory dotykowe i komórki linii bocznej ryb, które odbierają ruch wody względem ciała ryby oraz komórki reagujące na rozciąganie mięśni i ścięgien itp.
3) Chemoreceptory, komórki reagujące na określone substancje chemiczne; ich aktywność leży u podstaw pracy narządów węchu i smaku.
4) Termoreceptory, komórki odbierające temperaturę.
5) Elektroreceptory, komórki reagujące na pola elektryczne w środowisku.
Być może postawilibyśmy dziś te pięć typów receptorów w miejsce pięciu zmysłów opisanych przez Arystotelesa.
Rozważmy teraz na przykład jeden z rodzajów komórek receptorowych - fotoreceptory.
fotoreceptory
Fotoreceptory w siatkówce kręgowców to pręciki i czopki. Już w 1866 roku niemiecki anatom M. Schultz odkrył, że u ptaków dziennych siatkówki zawierają głównie czopki, podczas gdy ptaki nocne mają pręciki. Doszedł do wniosku, że pręciki służą do postrzegania słabego światła, a czopki - silnego. Wniosek ten potwierdziły późniejsze badania. Porównanie różnych zwierząt dodało wiele argumentów przemawiających za tą hipotezą: na przykład ryby głębinowe z ogromnymi oczami mają tylko pręciki w siatkówce.
Spójrz na ryc. 59. Przedstawia kij kręgowca. Ma segment wewnętrzny i segment zewnętrzny połączone szyjką. W obszarze segmentu wewnętrznego pręcik tworzy synapsy i uwalnia mediator, który oddziałuje na związane z nim neurony siatkówki. Mediator jest uwalniany, podobnie jak w innych komórkach, podczas depolaryzacji. W segmencie zewnętrznym znajdują się specjalne formacje - dyski, w których błonie osadzone są cząsteczki rodopsyny. Białko to jest bezpośrednim „odbiornikiem” światła.
Podczas badania pręcików okazało się, że pręcik może być wzbudzony zaledwie jednym fotonem światła, czyli ma najwyższą możliwą czułość. Kiedy jeden foton jest absorbowany, pole magnetyczne pręta zmienia się o około 1 mV. Z obliczeń wynika, że dla takiego przesunięcia potencjału konieczne jest oddziaływanie na około 1000 kanałów jonowych. Jak jeden foton może wpływać na tak wiele kanałów? Wiadomo było, że foton wnikając w pręcik jest wychwytywany przez cząsteczkę rodopsyny i zmienia stan tej cząsteczki.
Ale pojedyncza cząsteczka nie jest lepsza niż pojedynczy foton. Pozostało całkowicie niezrozumiałe, jak ta cząsteczka potrafi zmienić MT pręcika, zwłaszcza że krążki z rodopsyną nie są elektrycznie połączone z zewnętrzną błoną komórki.
Klucz do działania różdżek został znaleziony głównie w ciągu ostatnich kilku lat. Okazało się, że rodopsyna, pochłonąwszy kwant światła, nabiera na pewien czas właściwości katalizatora i udaje mu się zmienić kilka cząsteczek specjalnego białka, które z kolei powodują inne reakcje biochemiczne. Tak więc pracę pręta tłumaczy się zajściem reakcji łańcuchowej, która rozpoczyna się, gdy pochłonięty zostanie tylko jeden kwant światła i prowadzi do pojawienia się wewnątrz pręta tysięcy cząsteczek substancji, która może oddziaływać na kanały jonowe od wewnątrz komórka.
Co robi ten mediator wewnątrzkomórkowy? Okazuje się, że membrana wewnętrznego segmentu wędki jest dość powszechna - standardowa w swoich właściwościach: zawiera kanały K, które tworzą PP. Ale błona segmentu zewnętrznego jest niezwykła: zawiera tylko kanały Ka. W spoczynku są otwarte i choć nie jest ich zbyt wiele, to wystarczy, aby przepływający przez nie prąd zmniejszył MP, depolaryzując pręt. Tak więc mediator wewnątrzkomórkowy jest w stanie zamknąć część kanałów Ka, podczas gdy rezystancja obciążenia wzrasta, a MP również wzrasta, zbliżając się do potencjału równowagi potasu. W rezultacie pręt ulega hiperpolaryzacji pod wpływem światła.
Teraz poświęć chwilę, aby pomyśleć o tym, czego się właśnie nauczyłeś, a będziesz zaskoczony. Okazuje się, że nasze fotoreceptory uwalniają najwięcej mediatora w ciemności, ale oświetlone uwalniają go mniej, a im mniej, tym jaśniejsze światło. To niesamowite odkrycie zostało dokonane w 1968 roku. Yu.A. Trifonov z laboratorium A.L. Zadzwoń, kiedy niewiele było wiadomo o mechanizmie pałeczek.
A więc tutaj spotkaliśmy się z innym rodzajem kanałów – kanałami sterowanymi z wnętrza komórki.
Jeśli porównamy fotoreceptor kręgowca i bezkręgowca, zobaczymy, że ich praca ma wiele wspólnego: istnieje barwnik, taki jak rodopsyna; sygnał ze wzbudzonego pigmentu jest przekazywany do błony zewnętrznej za pomocą mediatora wewnątrzkomórkowego; komórka nie jest zdolna do generowania AP. Różnica polega na tym, że mediator wewnątrzkomórkowy działa w różnych organizmach na różne kanały jonowe: u kręgowców powoduje hiperpolaryzację receptora, podczas gdy u bezkręgowców z reguły powoduje depolaryzację. Na przykład u mięczaka morskiego – przegrzebka – gdy receptory dystalnej siatkówki są oświetlone, następuje ich hiperpolaryzacja, jak u kręgowców, ale jej mechanizm jest zupełnie inny. W przegrzebku światło zwiększa przepuszczalność błony dla jonów potasu, a MP przesuwa się bliżej równowagi potencjału potasu.
Jednak znak zmiany potencjału fotoreceptora nie jest zbyt duży, zawsze można go zmienić w toku dalszego przetwarzania. Ważne jest tylko, aby sygnał świetlny był niezawodnie przekształcany w sygnał elektryczny.
Rozważmy na przykład dalsze losy sygnału elektrycznego, który powstał w znanym nam układzie wzrokowym pąkli. U tych zwierząt fotoreceptory ulegają depolaryzacji po oświetleniu i uwalniają więcej przekaźnika, ale nie powoduje to żadnej reakcji zwierzęcia. Ale kiedy oczy są zacienione, rak zaczyna działać: usuwa czułki itp. Jak to się dzieje? Faktem jest, że mediator fotoreceptorów w pąkli jest hamujący, hiperpolaryzuje kolejną komórkę łańcucha neuronowego i zaczyna uwalniać mniej mediatora, więc gdy światło staje się jaśniejsze, nie zachodzi żadna reakcja. Przeciwnie, gdy fotoreceptor jest zacieniony, uwalnia mniej mediatora i przestaje hamować komórkę drugiego rzędu. Następnie ta komórka depolaryzuje i pobudza swoją komórkę docelową, w której powstają impulsy. Komórka 2 w tym obwodzie nazywana jest komórką I, od słowa „odwracająca”, ponieważ jej główną rolą jest zmiana znaku sygnału fotoreceptora. Pąkla ma raczej prymitywne oczy i potrzebuje trochę; prowadzi przywiązany tryb życia i wystarczy mu wiedzieć, że zbliża się wróg. U innych zwierząt system neuronów drugiego i trzeciego rzędu jest znacznie bardziej skomplikowany,
W fotoreceptorach potencjał receptora jest przekazywany dalej elektrotonicznie i wpływa na ilość uwalnianego mediatora. U kręgowców lub pąkli następna komórka jest pozbawiona impulsów i tylko trzeci neuron w łańcuchu jest w stanie generować impulsy. Ale w receptorze rozciągania naszych mięśni sytuacja jest zupełnie inna. Ten mechanoreceptor jest końcem włókna nerwowego, które owija się wokół włókna mięśniowego. Po rozciągnięciu cewki helisy utworzone przez niemielinową część włókna oddalają się od siebie i powstaje w nich potencjał receptora G - depolaryzacja z powodu otwarcia kanałów Ka wrażliwych na odkształcenie błony; potencjał ten wytwarza prąd przez punkt przecięcia Ranviera tego samego włókna, a punkt przecięcia generuje impulsy. Im bardziej mięsień jest rozciągnięty, tym większy potencjał receptorowy i wyższa częstotliwość impulsów.

W przypadku tego mechanoreceptora zarówno przekształcenie wpływu zewnętrznego w sygnał elektryczny, tj. w potencjał receptora, jak i przekształcenie potencjału receptora w impulsy są realizowane przez odcinek jednego aksonu.
Oczywiście interesujące byłoby dla nas omówienie budowy różnych receptorów u różnych zwierząt, ponieważ pod względem budowy i zastosowania są one bardzo egzotyczne; jednak każda taka historia sprowadzałaby się ostatecznie do tego samego: jak sygnał zewnętrzny jest przekształcany w potencjał receptora, który kontroluje uwalnianie neuroprzekaźnika lub powoduje generowanie impulsów.
Ale nadal będziemy mówić o jednym typie receptorów. To jest elektroreceptor. Jego osobliwość polega na tym, że sygnał, na który należy odpowiedzieć, ma już charakter elektryczny. Co robi ten receptor? Konwertuje sygnał elektryczny na elektryczny?
Elektroreceptory. Jak rekiny wykorzystują prawo i prawdopodobieństwo Ohma
w 1951 roku angielski naukowiec Lissman badał zachowanie ryb hymnarch. Ryba ta żyje w błotnistych, nieprzejrzystych wodach w jeziorach i bagnach Afryki i dlatego nie zawsze może używać wzroku do orientacji. Lissman zasugerował, że te ryby, podobnie jak nietoperze, używają echolokacji do orientacji.
Niesamowitą zdolność nietoperzy do latania w całkowitej ciemności bez wpadania na przeszkody odkryto bardzo dawno temu, w 1793 roku, czyli niemal równocześnie z odkryciem Galvaniego. Dokonał tego Lazaro Spallanzani, profesor Uniwersytetu w Pawii. Jednak eksperymentalny dowód na to, że nietoperze emitują ultradźwięki i poruszają się za pomocą ich echa, uzyskano dopiero w 1938 roku na Uniwersytecie Harvarda w USA, kiedy to fizycy stworzyli sprzęt do rejestracji ultradźwięków.
Po eksperymentalnym przetestowaniu hipotezy orientacji hymnarchy ultradźwiękowej, Lissman ją odrzucił. Okazało się, że hymnarcha był prowadzony jakoś inaczej. Badając zachowanie hymnarchy, Lissman odkrył, że ta ryba ma narząd elektryczny iw nieprzejrzystej wodzie zaczyna generować wyładowania o bardzo słabym prądzie. Taki prąd nie nadaje się ani do obrony, ani do ataku. Następnie Lissman zasugerował, że hymnarcha powinien mieć specjalne narządy do postrzegania pól elektrycznych - system elektrosensoryczny.
To była bardzo odważna hipoteza. Naukowcy wiedzieli, że owady widzą światło ultrafioletowe, a wiele zwierząt słyszy dźwięki, które są dla nas niesłyszalne. Było to jednak tylko pewne poszerzenie zakresu percepcji sygnałów, które ludzie również mogą odbierać. Lissman dopuścił istnienie zupełnie nowego typu receptora.
Sytuację komplikował fakt, że znana była już wówczas reakcja ryb na słabe prądy. Została zaobserwowana w 1917 roku przez Parkera i Van Heusera na sumie. Jednak autorzy ci podali swoje obserwacje zupełnie inne wyjaśnienie. Uznali, że gdy prąd przepływa przez wodę, zmienia się w niej rozkład jonów, a to wpływa na smak wody. Ten punkt widzenia wydawał się całkiem prawdopodobny: po co wymyślać nowe narządy, skoro wyniki można wytłumaczyć znanymi zwykłymi narządami smaku. To prawda, że naukowcy ci w żaden sposób nie udowodnili swojej interpretacji; nie zorganizowali eksperymentu kontrolnego. Gdyby odcięli nerwy z narządów smaku, tak że wrażenia smakowe u ryb zniknęły, odkryliby, że reakcja na prąd została zachowana. Ograniczając się do werbalnego wyjaśnienia swoich obserwacji, przegapili wielkie odkrycie.
Z drugiej strony Lissman wymyślił i przeprowadził wiele różnych eksperymentów i po dziesięciu latach pracy udowodnił swoją hipotezę. Około 25 lat temu nauka uznała istnienie elektroreceptorów. Zaczęto badać elektroreceptory i wkrótce znaleziono je u wielu ryb morskich i słodkowodnych, a także u minogów. Około 5 lat temu odkryto takie receptory u płazów, a ostatnio u ssaków.
Gdzie znajdują się elektroreceptory i jak są rozmieszczone?
Ryby mają mechanoreceptory linii bocznej rozmieszczone wzdłuż ciała i na głowie ryby; dostrzegają ruch wody względem zwierzęcia. Elektroreceptory to kolejny typ receptora linii bocznej. Podczas rozwoju embrionalnego wszystkie receptory linii bocznej rozwijają się z tego samego regionu układu nerwowego, co receptory słuchowe i przedsionkowe. Tak więc receptory słuchowe nietoperzy i elektroreceptory ryb są bliskimi krewnymi.
U różnych ryb elektroreceptory mają różną lokalizację - znajdują się na głowie, na płetwach, wzdłuż ciała, a także mają inną strukturę. Często komórki elektroreceptorowe tworzą wyspecjalizowane narządy. Przyjrzymy się tutaj jednemu z tych narządów występujących u rekinów i płaszczek, brodawce Lorenziniego. Lorenzini uważał, że ampułki to gruczoły wytwarzające rybi śluz. Brodawka Lorenziniego jest kanałem podskórnym, którego jeden koniec jest otwarty na środowisko zewnętrzne, a drugi kończy się głuchym przedłużeniem; światło kanału jest wypełnione galaretowatą masą; komórki elektroreceptorowe wyściełają „dno” ampułki w jednym rzędzie.
Ciekawe, że Parker, który jako pierwszy zauważył, że ryby reagują na słabe prądy elektryczne, badał również ampułki Lorenziniego, ale przypisywał im zupełnie inne funkcje. Odkrył, że przyciskając patyk do zewnętrznego wejścia do kanału, rekin może zareagować. Na podstawie takich eksperymentów wywnioskował, że ampułka Lorenziniego była manometrem do pomiaru głębokości zanurzenia ryb, zwłaszcza że organ miał budowę podobną do manometru. Ale i tym razem interpretacja Parkera była błędna. Jeśli rekin zostanie umieszczony w komorze ciśnieniowej i powstanie w niej podwyższone ciśnienie, to ampułka Lorenziniego na to nie zareaguje - a tego nie widać bez eksperymentowania: woda naciska ze wszystkich stron i nie ma efektu*). A przy ciśnieniu tylko na por w galarecie, która go wypełnia, powstaje różnica potencjałów, tak jak powstaje różnica potencjałów w krysztale piezoelektrycznym.
Jak ułożone są ampułki Lorenziniego? Okazało się, że wszystkie komórki nabłonka wyściełającego kanał są mocno połączone specjalnymi „ciasnymi stykami”, co zapewnia wysoką oporność właściwą nabłonka. Kanał pokryty tak dobrą izolacją przechodzi pod skórą i może mieć kilkadziesiąt centymetrów długości. Natomiast galaretka wypełniająca kanał brodawki Lorenziniego ma bardzo niską rezystywność; zapewnia to fakt, że pompy jonowe pompują wiele jonów K+ do światła kanału. Kanał organów elektrycznych to więc kawałek dobrego kabla o wysokiej rezystancji izolacji i dobrze przewodzącym rdzeniu.
„Dno” ampułki pokrywa w jednej warstwie kilkadziesiąt tysięcy komórek elektroreceptorowych, które również są mocno sklejone. Okazuje się, że komórka receptorowa patrzy z jednej strony do wnętrza kanału, a na drugim tworzy synapsę, gdzie uwalnia mediator pobudzający, który oddziałuje na zbliżające się do niej zakończenie włókna nerwowego. Każda ampułka ma 10-20 włókien aferentnych, a każda z nich daje wiele zakończeń, które prowadzą do receptorów, tak że około 2000 komórek receptorowych działa na każde włókno.
Zobaczmy teraz, co dzieje się z samymi komórkami elektroreceptorowymi pod działaniem pola elektrycznego.
Jeśli dowolne ogniwo zostanie umieszczone w polu elektrycznym, to w jednej części membrany znak GSH będzie się pokrywał ze znakiem natężenia pola, aw drugiej części okaże się, że jest przeciwny. Oznacza to, że na jednej połowie komórki MP wzrośnie, a na drugiej przeciwnie, zmniejszy się. Okazuje się, że każda komórka „czuje” pola elektryczne, czyli jest elektroreceptorem.

I to jest zrozumiałe: wszak w tym przypadku znika problem zamiany sygnału zewnętrznego na sygnał naturalny dla komórki – elektryczny. Tak więc komórki elektroreceptorowe działają w bardzo prosty sposób: przy odpowiednim znaku pola zewnętrznego błona synaptyczna tych komórek ulega depolaryzacji, a to przesunięcie potencjału steruje uwalnianiem mediatora.
Ale wtedy pojawia się pytanie: jakie są cechy komórek elektroreceptorowych? Czy każdy neuron może wykonywać swoje funkcje? Jaki jest specjalny układ ampułek Lorenziniego?
Tak, jakościowo każdy neuron można uznać za elektroreceptor, ale jeśli przejdziemy do szacunków ilościowych, sytuacja się zmieni. Naturalne pola elektryczne są bardzo słabe, a wszelkie sztuczki, jakie natura stosuje w narządach elektroczułych mają na celu po pierwsze wyłapanie jak największej różnicy potencjałów na błonie synaptycznej, a po drugie zapewnienie wysokiej czułości mechanizmu uwalniania mediatora na zmiany MP.
Narządy elektryczne rekinów i płaszczek są niezwykle wrażliwe: ryby reagują na pola elektryczne o natężeniu 0,1 μV/cm. Tak więc problem wrażliwości został w przyrodzie znakomicie rozwiązany. Jak osiąga się takie wyniki?
Po pierwsze, budowa ampułki Lorenziniego przyczynia się do zapewnienia takiej wrażliwości. Jeśli natężenie pola wynosi 0,1 μV/cm, a długość kanału ampułki wynosi 10 cm, to cała ampułka będzie miała różnicę potencjałów 1 μV. Prawie całe to napięcie spadnie na warstwę odbiorczą, ponieważ jej rezystancja jest znacznie wyższa niż rezystancja ośrodka w kanale. Rekin tutaj bezpośrednio korzysta z prawa Ohma: V \u003d 11 $, ponieważ prąd płynący w obwodzie jest taki sam, spadek napięcia jest większy tam, gdzie rezystancja jest wyższa. Zatem im dłuższy kanał ampułki i im mniejszy opór, tym większa różnica potencjałów jest przykładana do elektroreceptora.
Po drugie, prawo Ohma jest „stosowane” przez same elektroreceptory; różne odcinki ich błony mają również różne opory: błona synaptyczna, w której uwalniany jest mediator, ma duży opór, a przeciwległa część błony ma mały, tak że tutaj również różnica potencjałów rozkłada się tak korzystnie jak to możliwe,
Jeśli chodzi o wrażliwość błony synaptycznej na przesunięcia w MF, można to wytłumaczyć różnymi przyczynami: kanały Ca tej błony lub sam mechanizm uwalniania mediatora mogą być bardzo wrażliwe na potencjalne przesunięcia. Bardzo ciekawe wyjaśnienie dużej wrażliwości uwalniania neuroprzekaźników na przesunięcia MP zaproponował A.L. Dzwonić. Jego pomysł polega na tym, że w takich synapsach prąd generowany przez błonę postsynaptyczną wpływa do komórek receptorowych i sprzyja uwalnianiu mediatora; w rezultacie następuje pozytywne sprzężenie zwrotne: uwolnienie neuroprzekaźnika powoduje PSP, podczas gdy prąd przepływa przez synapsę, co wzmaga uwalnianie neuroprzekaźnika. W zasadzie taki mechanizm musi koniecznie działać. Ale nawet w tym przypadku pytanie jest ilościowe: jak skuteczny jest taki mechanizm, aby odgrywać jakąkolwiek rolę funkcjonalną? Ostatnio A.L. Callowi i jego współpracownikom udało się uzyskać przekonujące dane eksperymentalne potwierdzające, że taki mechanizm rzeczywiście działa w fotoreceptorach.
Kontrola hałasu
Tak więc, dzięki różnym sztuczkom wykorzystującym prawo Ohma, na błonie elektroreceptorów powstaje potencjalne przesunięcie rzędu 1 μV. Wydawać by się mogło, że jeśli czułość błony presynaptycznej jest wystarczająco wysoka – a tak jest, jak widzieliśmy – to wszystko jest w porządku. Ale nie wzięliśmy pod uwagę, że zwiększenie czułości dowolnego urządzenia rodzi nowy problem - problem kontroli hałasu. Nazwaliśmy czułość elektroreceptora, który odbiera 1 μV, fantastyczną, a teraz wyjaśnimy, dlaczego. Faktem jest, że ta wartość jest znacznie niższa niż poziom hałasu.
W każdym przewodniku nośniki ładunku uczestniczą w ruchu termicznym, tj. poruszają się losowo w różnych kierunkach. Czasami więcej ładunków porusza się w jednym kierunku niż w drugim, co oznacza, że w dowolnym przewodniku bez źródła e. ds. występują prądy. W odniesieniu do metali problem ten rozważali już w 1913 roku de Haas i Lorentz. Eksperymentalnie szum termiczny w przewodnikach został odkryty w 1927 roku przez Johnsona. W tym samym roku G. Nyquist podał szczegółową i ogólną teorię tego zjawiska. Teoria i eksperyment były zgodne: wykazano, że natężenie szumu zależy liniowo od wartości rezystancji i temperatury przewodnika. Jest to naturalne: im większy opór przewodnika, tym większa różnica potencjałów, która pojawia się na nim z powodu przypadkowych prądów, a im wyższa temperatura, tym większa prędkość nośników ładunku. Zatem im większy opór przewodnika, tym większe są w nim fluktuacje potencjału pod wpływem ruchu termicznego ładunków.
A teraz wróćmy do elektroreceptorów. Powiedzieliśmy, że aby zwiększyć czułość w tym odbiorniku, korzystnie jest mieć jak największą rezystancję membrany, tak aby większość napięcia na niej spadała. Rzeczywiście, opór błony uwalniającej mediator jest bardzo wysoki w komórce elektroreceptorowej, rzędu 10 10 omów. Jednak wszystko ma swoją cenę: wysoka odporność tej membrany prowadzi do zwiększonego hałasu. Fluktuacja potencjału na membranie elektroreceptora spowodowana szumem termicznym wynosi około 30 μV, tj. 30 razy więcej niż minimalne postrzegane przesunięcie MF, które występuje pod działaniem pola zewnętrznego! Okazuje się, że sytuacja wygląda tak, jakbyś siedział w pokoju, w którym każda z trzech tuzinów osób mówi o sobie, a ty próbujesz nawiązać rozmowę z jedną z nich. Jeśli głośność wszystkich dźwięków jest 30 razy głośniejsza niż głośność twojego głosu, rozmowa będzie oczywiście niemożliwa.
Jak rekin „słyszy” taką rozmowę przez szum termiczny? Czy mamy do czynienia z cudem? Oczywiście nie. Poprosiliśmy o zwrócenie uwagi na fakt, że na jednym włóknie czuciowym synapsuje około 2000 elektroreceptorów. Pod wpływem szumów termicznych w błonie neuroprzekaźnik jest uwalniany z jednej lub drugiej synapsy, a włókno doprowadzające, nawet przy braku pól elektrycznych na zewnątrz ryby, stale impulsuje. Kiedy pojawia się sygnał zewnętrzny, wszystkie 2000 komórek wydziela mediator, w wyniku czego sygnał zewnętrzny jest wzmacniany.
Czekaj, powie myślący czytelnik, bo 2000 komórek powinno robić więcej hałasu! Okazuje się, że jeśli kontynuować analogię z rozmową w hałaśliwym pokoju, to 100 osób łatwiej przekrzycze trzytysięczny tłum niż jeden - trzydzieści? Ale okazuje się, że w rzeczywistości, co dziwne, jest tak, jak jest. Zapewne każdy z nas nie raz słyszał, jak rytmiczne, nasilające się oklaski przebijają się przez burzę oklasków. Lub poprzez ryk trybun stadionu wyraźnie słychać okrzyki: „Brawo! Dobra robota!”, skandowane nawet przez niewielką grupę fanów. Faktem jest, że we wszystkich tych przypadkach mamy do czynienia z konfrontacją zorganizowanego, synchronicznego sygnału z szumem, czyli sygnałem chaotycznym. Z grubsza mówiąc, wracając do elektroreceptorów, ich reakcje na sygnał zewnętrzny są synchroniczne i sumują się, a tylko część losowego szumu termicznego pokrywa się w czasie. Dlatego amplituda sygnału rośnie wprost proporcjonalnie do liczby komórek receptorowych, podczas gdy amplituda szumu rośnie znacznie wolniej. Ale niech czytelnik interweniuje jeszcze raz, skoro szum w odbiorniku jest tylko 30 razy silniejszy niż sygnał, czy natura nie jest zbyt marnotrawna? Po co 2000 receptorów? Może wystarczy sto?
Jeśli chodzi o problemy ilościowe, musisz liczyć, co oznacza, że potrzebujesz matematyki. W matematyce istnieje specjalna sekcja - teoria prawdopodobieństwa, w której badane są losowe zjawiska i procesy o bardzo różnym charakterze. Niestety ten dział matematyki w ogóle nie jest wprowadzany w szkole ogólnokształcącej.
Teraz zróbmy proste obliczenie. Niech pole zewnętrzne przesunie MP wszystkich receptorów o 1 μV, wtedy całkowity użyteczny sygnał wszystkich receptorów będzie równy 2000 pewnych jednostek. Średnia wartość sygnału szumu jednego receptora wynosi około 30 μV, ale całkowity sygnał szumu jest proporcjonalny do 2000, czyli równy tylko 1350 jednostkom. Widzimy, że dzięki zsumowaniu efektu dużej liczby receptorów użyteczny sygnał jest 1,5 razy większy niż szum. Widać, że nie można zrezygnować ze stu komórek receptorowych. A przy stosunku sygnału do szumu wynoszącym 1,5 układ nerwowy rekina jest już w stanie wykryć ten sygnał, więc cud się nie dzieje.
Powiedzieliśmy, że pręciki siatkówki reagują na wzbudzenie tylko jednej cząsteczki rodopsyny. Ale takie wzbudzenie może powstać nie tylko pod wpływem światła, ale także pod wpływem szumu termicznego. Ze względu na dużą czułość pręcików w siatkówce zawsze powinny pojawiać się sygnały „fałszywego alarmu”. Jednak w rzeczywistości siatkówka ma również system kontroli hałasu oparty na tej samej zasadzie. Pręciki są ze sobą połączone przez ES, co prowadzi do uśrednienia ich potencjalnych przesunięć, dzięki czemu wszystko dzieje się tak samo, jak w elektroreceptorach. Należy również pamiętać o skojarzeniu poprzez wysoce przepuszczalne styki spontanicznie aktywnych komórek węzła zatokowego serca, co daje regularny rytm serca i eliminuje fluktuacje właściwe dla pojedynczej komórki. Widzimy, że natura szeroko wykorzystuje uśrednianie, aby radzić sobie z hałasem w różnych sytuacjach.
Jak zwierzęta wykorzystują swoje elektroreceptory? W przyszłości porozmawiamy bardziej szczegółowo o metodzie orientacji ryb w błotnistej wodzie. Ale rekiny i płaszczki używają swoich elektroreceptorów podczas poszukiwania zdobyczy. Drapieżniki te są w stanie wykryć flądrę ukrytą pod warstwą piasku jedynie dzięki polu elektrycznemu generowanemu przez jej mięśnie podczas ruchów oddechowych. Ta zdolność rekinów została pokazana w serii pięknych eksperymentów przeprowadzonych przez Kelmina w 1971 roku. Zwierzę może leżeć nisko i się nie ruszać, może maskować się jako kolor tła, ale nie może zatrzymać metabolizmu, zatrzymać bicia serca, przestać oddychać, więc zapachy zawsze ją demaskują, aw wodzie - i pola elektryczne powstające w wyniku pracy serca i innych mięśni. Tak wiele drapieżnych ryb można nazwać „elektrycznymi detektywami”.

... ; przeciwciała IgG4, IgA, IgD i IgE nie aktywują dopełniacza. Do funkcji efektorowych immunoglobulin należy również ich selektywne oddziaływanie z różnymi typami komórek przy udziale specjalnych receptorów powierzchniowych komórki. RECEPTORY KOMÓRKOWE PRZECIWCIAŁ Istnieją trzy typy receptorów powierzchniowych komórek dla IgG Receptory komórkowe dla IgG pośredniczą w szeregu funkcji efektorowych...
1. Centralny układ nerwowy
Centralny układ nerwowy jest częścią układu nerwowego kręgowców, reprezentowaną przez nagromadzenie komórek nerwowych tworzących mózg i rdzeń kręgowy.
Centralny układ nerwowy reguluje procesy zachodzące w organizmie i pełni funkcję centrum sterowania wszystkimi układami. Mechanizmy działania ośrodkowego układu nerwowego opierają się na wzajemnym oddziaływaniu pobudzenia i hamowania.
Wyższa aktywność nerwowa (HNI)
Wyższa aktywność nerwowa - według I.P. Pavlova - jest złożoną formą aktywności życiowej, która zapewnia indywidualną adaptację behawioralną ludzi i zwierząt wyższych do zmieniających się warunków środowiskowych.
Wyższa aktywność nerwowa opiera się na interakcji wrodzonych odruchów bezwarunkowych z odruchami warunkowymi nabytymi w procesie ontogenezy, do których u człowieka dodawany jest drugi układ sygnalizacyjny.
Podstawą strukturalną VND jest kora mózgowa z jądrami podkorowymi przodomózgowia i niektórymi strukturami międzymózgowia.
2. Wyższa aktywność nerwowa
Wyższa aktywność nerwowa (HNA) to aktywność wyższych działów ośrodkowego układu nerwowego, która zapewnia najdoskonalsze przystosowanie zwierząt i ludzi do środowiska (zachowanie). Podstawą strukturalną DNB jest kora mózgowa z jądrami podkorowymi przodomózgowia i formacjami międzymózgowia, jednak nie ma sztywnego związku między DNB a strukturami mózgu. Niższa aktywność nerwowa została przedstawiona jako funkcja ośrodkowego układu nerwowego, mająca na celu regulację procesów fizjologicznych w samym organizmie. Najważniejszą cechą DNB jest charakter sygnałowy, który pozwala zawczasu przygotować się do takiej czy innej formy aktywności (żywnościowej, obronnej, seksualnej itp.)
Charakterystyka DNB: zmienność, sygnalizacja, adaptacyjność - zapewniają elastyczność i adaptacyjność reakcji. Probabilistyczny charakter środowiska zewnętrznego daje względność każdej reakcji behawioralnej i skłania organizm do prognozowania probabilistycznego. Zdolność uczenia się w dużym stopniu zależy nie tylko od procesów pobudzenia, ale także hamowania. Hamowanie warunkowe przyczynia się do szybkiej zmiany zachowania w zależności od warunków i motywacji.
Termin DNB został wprowadzony przez I.P. Pavlova, który uznał go za równoważny z pojęciem „aktywności umysłowej”. Według IP Pavlova jest to połączona funkcja odruchowa (odruch warunkowy i bezwarunkowy) kory mózgowej i najbliższej podkory mózgowej. Wprowadził również pojęcie „systemów sygnałowych” jako systemu warunkowych połączeń odruchowych, podkreślając pierwszy system sygnałowy wspólny dla zwierząt i ludzi, a drugi specyficzny tylko dla ludzi.
Pierwszy system sygnalizacyjny (PSS) - bezpośrednie doznania i spostrzeżenia, stanowi podstawę DNB i sprowadza się do zestawu różnorodnych odruchów warunkowych i bezwarunkowych na bezpośrednie bodźce. Ludzki PSS charakteryzuje się większą szybkością rozprzestrzeniania się i koncentracji procesu nerwowego, jego ruchliwością, co zapewnia szybkość przełączania i powstawanie odruchów warunkowych. Zwierzęta lepiej rozróżniają poszczególne bodźce, człowieka – ich kombinacje.
Drugi system sygnałowy ukształtował się u człowieka na bazie pierwszego jako system sygnałów mowy (wymawiane, słyszalne, widoczne). Słowa zawierają uogólnienie sygnałów pierwszego systemu sygnałowego. Proces uogólniania przez słowo rozwija się w trakcie tworzenia odruchów warunkowych. Uogólniona refleksja i abstrakcja powstają dopiero w procesie komunikacji, tj. determinowane czynnikami biologicznymi i społecznymi.
Receptor - (od łac. recepturre - otrzymywać), formacje nerwowe, które przekształcają chemiczne i fizyczne wpływy z zewnętrznego lub wewnętrznego środowiska ciała na impulsy nerwowe; obwodowa wyspecjalizowana część analizatora, za pomocą której tylko określony rodzaj energii jest przekształcany w proces pobudzenia nerwowego. Receptory różnią się znacznie stopniem złożoności strukturalnej i poziomem przystosowania do pełnionych funkcji. W zależności od energii odpowiedniej stymulacji, receptory dzielą się na mechanoreceptory i chemoreceptory. Mechanoreceptory znajdują się w uchu, aparacie przedsionkowym, mięśniach, stawach, skórze i narządach wewnętrznych. Chemoreceptory służą zmysłowi węchu i smaku: wiele z nich znajduje się w mózgu, reagując na zmiany składu chemicznego płynnego środowiska organizmu. Receptory wzrokowe są również zasadniczo chemoreceptorami. W zależności od umiejscowienia w ciele i pełnionej funkcji, receptory dzielą się na eksteroreceptory, interoreceptory i proprioreceptory. Eksteroreceptory obejmują odległe receptory, które odbierają informacje w pewnej odległości od źródła podrażnienia (węchowe, słuchowe, wzrokowe, smakowe); interoreceptory informują o bodźcach ze środowiska wewnętrznego, a proprioreceptory o stanie narządu ruchu organizmu. Poszczególne receptory są ze sobą anatomicznie połączone i tworzą pola receptywne, które mogą się nakładać.
3. Receptor
Od łac. Receptum - przyjąć
Receptor – wrażliwe zakończenie nerwowe lub wyspecjalizowana komórka, która przetwarza odczuwane podrażnienie na impulsy nerwowe.
Wszystkie receptory charakteryzują się obecnością określonego odcinka błony zawierającego białko receptorowe, które determinuje procesy odbioru. W zależności od wybranej klasyfikacji receptory dzielą się na:
Na poziomie podstawowym i wtórnym;
Na foto-, phono-, termo-, elektro- i baro-;
Na zewnątrz- i intero-;
Na mechano-, foto- i chemo-;
Na nocyreceptorach, ciepło, zimno, dotyk itp.;
Na mono- i wielowartościowych;
Słuchowe, wizualne, węchowe, dotykowe i smakowe;
Do kontaktu i na odległość;
Na fazowe, toniczne i fazowo-toniczne.
Rodzaje receptorów. Adaptacja mechanizmów receptorowych
Adaptacja mechanizmów receptorowych - proces zmniejszania (zmniejszania) aktywności receptorów jako bodźca o stałej charakterystyce fizycznej.
Charakter adaptacji mechanizmów receptorowych zależy od:
Z właściwości aparatu pomocniczego;
Z cech struktur percepcyjnych receptora;
Z właściwości elementów regeneracyjnych zakończenia nerwowego;
Dla wtórnych receptorów czuciowych: od właściwości kontaktu synaptycznego między komórką receptorową a końcem neuronu czuciowego.
receptor bólu
nocyreceptor; nocyceptor
Receptor bólu to receptor, który jest podrażniony, aby spowodować ból.
przedsionkowo-receptorowe
Akceleroreceptory
Vestibuloreceptory - receptory, które dostrzegają zmiany prędkości i kierunku ruchu ciała w przestrzeni. U ludzi przedsionkowo-receptory są reprezentowane przez komórki rzęsate błoniastego błędnika ucha wewnętrznego.
kubki smakowe
Receptory smaku - chemoreceptory, których podrażnienie powoduje odczucia smakowe.
Kubki smakowe:
Zlokalizowane w błonie śluzowej jamy ustnej;
Reagują na cztery rodzaje substancji: kwaśne, słone, gorzkie i słodkie.
Wtórny receptor czuciowy
Receptor niewolny
Receptor wtórnego wykrywania - receptor, który jest wyspecjalizowaną komórką, której wzbudzenie jest przekazywane do zakończeń odpowiedniego neuronu doprowadzającego.
Glukoreceptory
Glukoreceptory to receptory wrażliwe na zmiany stężenia glukozy we krwi.
Odległy receptor
telereceptor
Odległy receptor - receptor odbierający bodźce, których źródło znajduje się w pewnej odległości od ciała.
Wizualne guzki
Wizualne pagórki są częścią międzymózgowia; główne podkorowe ośrodki wrażliwości. Impulsy ze wszystkich receptorów ciała wchodzą do guzków wzrokowych wzdłuż wznoszących się ścieżek, a stąd do kory mózgowej.
Interoreceptor
Interoceptor; wisceroreceptor; Receptor wewnętrzny
Od łac. Wnętrze - wewnętrzne + Capio - weź
Interoreceptor - receptor:
Zlokalizowane w narządach wewnętrznych, tkankach lub naczyniach; I
Dostrzeganie zmian mechanicznych, chemicznych i innych w środowisku wewnętrznym organizmu.
Receptor skóry
Receptor skórny – receptor znajdujący się w skórze i zapewniający odczuwanie podrażnienia mechanicznego, temperaturowego i bólowego.
Mechanoreceptor
Mechanoreceptor - wrażliwe zakończenie nerwowe, które odbiera wpływy mechaniczne: nacisk, przyspieszenie itp.
Receptor monomodalny
Receptor monowalentny
Receptor monomodalny - receptor odbierający tylko jeden rodzaj podrażnienia.
Receptory węchowe
Receptory węchowe - chemoreceptory błony śluzowej górnych partii jamy nosowej, których podrażnienie powoduje węch.
Pierwotny receptor czuciowy
Pierwotny receptor czuciowy - receptor będący wrażliwym zakończeniem nerwowym.
Receptor polimodalny
Receptor wielowartościowy
Receptor polimodalny - receptor, który odbiera kilka rodzajów bodźców.
receptory tkankowe
Receptory tkankowe - receptory zlokalizowane w narządach i tkankach poza wyspecjalizowanymi strefami refleksogennymi.
Receptor toniczny
Receptor toniczny - termoreceptor, pręcik siatkówkowy lub inny wolno adaptujący się receptor, który reaguje w mniej lub bardziej stały sposób na bezwzględną wielkość bodźca.
Chemoreceptory
Chemoceptory; Chemoreceptory
Chemoreceptory to wyspecjalizowane wrażliwe komórki lub struktury komórkowe, przez które organizm zwierząt i ludzi odbiera bodźce chemiczne, w tym zmiany w metabolizmie. Oddziaływanie chemikaliów na chemoreceptory prowadzi do pojawienia się potencjałów bioelektrycznych w chemoreceptorach.
Zewnętrzny receptor
eksteroreceptor; Receptor zewnętrzny
Od łac. Exter - lat + Recipere - weź
Exteroreceptor – receptor zlokalizowany na powierzchni ciała i odbierający podrażnienia pochodzące ze środowiska zewnętrznego. Zazwyczaj eksteroreceptory są wyspecjalizowanymi formacjami nabłonka nerwowego.
Receptor jest narządem roboczym obwodowej części wrażliwego neuronu. Ciało neuronu znajduje się w zwoju międzykręgowym. Wyrostek obwodowy zwoju pseudojednobiegunowego kończy się w tkankach z receptorem, natomiast wyrostek ośrodkowy wchodzi do rdzenia kręgowego i uczestniczy w tworzeniu różnych dróg czuciowych.
Włókna nerwów czuciowych są podzielone na gałęzie, które są wysyłane do różnych części tej samej tkanki lub do kilku różnych tkanek. Zakończenia nerwowe - receptory - mogą znajdować się bezpośrednio na strukturach roboczych otaczających tkanek, w takich przypadkach nazywane są wolnymi. Inne sąsiadują z powierzchnią specjalnych komórek pomocniczych i tworzą niewolne zakończenia. Niewolne zakończenia mogą być zamknięte w mniej lub bardziej złożonej kapsułce składającej się z komórek pomocniczych (otoczonych receptorów). Według histologów komórki pomocnicze pełnią funkcje tkanki podporowej i uczestniczą w procesie pobudzenia.
Z punktu widzenia specjalizacji funkcjonalnej zwyczajowo rozróżnia się ekstero-, proprio- i interoreceptory. Eksteroreceptory, jak sama nazwa wskazuje, znajdują się na tkankach powłokowych człowieka i są w większości reprezentowane przez wolne zakończenia. Niektóre włókna nerwowe silnie rozgałęziają się i tworzą krzaki, których gałęzie kończą się włóknistymi siatkami lub zgrubieniami między komórkami nabłonka, podczas gdy inne wychodzą na wolną powierzchnię nabłonka bez rozgałęzień, a nawet wychodzą na jego powierzchnię. Końcowe odcinki takich receptorów wraz z złuszczającymi się komórkami nabłonka obumierają i odrywają się, co wyraża się zwiększoną aktywnością regeneracyjną receptorów o tej strukturze. Spośród wyspecjalizowanych receptorów tkanek powłokowych należy wymienić niewolne zakończenia znajdujące się w narządach smaku (kubki smakowe, opuszki itp.), ciała dotykowe Merkel, opuszki węchowe itp. części ciała (przegroda nosowa) .
Głębsze receptory znajdują się w mięśniach, powięziach, więzadłach, okostnej, naczyniach i nerwach.
Receptor dla tkanki mięśni poprzecznie prążkowanych jest wyspecjalizowaną formacją wrzeciona nerwowo-mięśniowego. Jest częścią jednego, dwóch lub trzech włókien mięśniowych o długości do kilku milimetrów, splecionych z gałązkami wrażliwego włókna nerwowego, które tworzy swoisty splot wokół włókien mięśniowych. Receptory te są wolne, reagując na rozciąganie tkanki mięśniowej.
Receptory mięśnia sercowego są reprezentowane przez wspomniane wrzeciona mięśniowe i „wspinające się” zakończenia nerwowe, zakończone szerokimi płytkami włóknistymi.
W mięśniach gładkich różnych narządów wewnętrznych znaleziono tylko krzaczaste receptory o różnych kształtach.
Najbardziej zróżnicowane są receptory tkanki łącznej i naczyniowe. Wśród nich wyróżnia się zakończenia wolne, niewolne i enkapsulowane. Częściej niż inne, w tkance łącznej wykrywa się różne krzaczaste lub przypominające drzewa receptory o różnym stopniu złożoności. Charakterystyczną formą receptorów tkanki łącznej są zakończenia nerwowe w postaci „kłębuszka”. Najluźniejsze „kulki” są przesiąknięte włóknami tkanki łącznej i są receptorami rozciągania, podczas gdy inne są stosunkowo odizolowane od otaczających tkanek, działając jako receptory nacisku. Istnieją również bardziej złożone zakończenia nerwowe w postaci ciałek Vatera-Pacchiniego, kolb Krause, Golgiego-Mazzoniego, ciałek Meissnera. Ustalono, że ciała Vatera-Pacchiniego są receptorami nacisku mechanicznego, kolby Krause temperatury, Golgiego-Mazzoniego nacisku i rozciągania, a Meissnera bodźców dotykowych.
Receptory naczyniowe są nie mniej zróżnicowane. Naczynia mają obfite wrażliwe unerwienie na całej długości od serca do wewnątrzorganicznych naczyń włosowatych. Główną formą receptorów są końcówki krzaczaste, które mogą być wolne i niewolne. Rejestrują stan rozciągnięcia ściany naczynia, wysokość ciśnienia krwi w naczyniach, skład chemiczny krwi. Charakterystyczną cechą receptorów naczyń wewnątrzorganicznych jest to, że pokrywają one swoimi odgałęzieniami obszar otaczającej tkanki (receptory tkanki naczyniowej). Receptory naczyń limfatycznych badano w mniejszym stopniu, są one reprezentowane przez zwykłe receptory tkanki łącznej.
Receptory obwodowego układu nerwowego i zwojów autonomicznych mają zróżnicowany kształt i pełnią funkcje ogólnego odbioru.
Impuls nerwowy generowany w receptorach potencjału czynnościowego włókien czuciowych dociera do pierwszej stacji przekaźnikowej do przetwarzania (percepcji) przepływu doprowadzającego w ośrodkowym układzie nerwowym. Rdzeń kręgowy (medulla spinalis) u dorosłych to pasmo o długości 41–45 cm, nieco spłaszczone od przodu do tyłu. Posiada dwa zgrubienia odpowiadające korzeniom nerwowym kończyny górnej i dolnej. Spośród tych zgrubień częściej spotyka się zgrubienie lędźwiowe, ale bardziej zróżnicowane jest szyjne, co wiąże się ze skomplikowaną zorganizowaną motoryką ręki. Funkcjonalnie należy podkreślić, że tej głównej funkcji podporządkowana jest organizacja kompleksów czuciowych na poziomie odcinków szyjnych.
Receptory (łac. receptor - akceptujący, od recipio - akceptuję, odbieram), specjalne wrażliwe formacje, które postrzegają i przekształcają podrażnienia z zewnętrznego lub wewnętrznego środowiska organizmu i przekazują informacje o substancji czynnej do układu nerwowego, receptora. charakteryzuje się różnorodnością pod względem konstrukcyjnym i funkcjonalnym. Mogą być reprezentowane przez wolne zakończenia włókien nerwowych, zakończenia pokryte specjalną torebką, a także wyspecjalizowane komórki w kompleksowo zorganizowanych formacjach, takich jak siatkówka, narząd Cortiego itp., składające się z wielu receptorów.
Chwytnik(od łacińskiego słowa - odbieranie) w biologii ma dwa znaczenia. W pierwszym sensie receptory nazywane są wrażliwymi zakończeniami nerwowymi lub wyspecjalizowanymi komórkami, które odbierają podrażnienia ze środowiska zewnętrznego lub wewnętrznego i przekształcają je w pobudzenie nerwowe, przekazywane w postaci strumienia impulsów nerwowych do ośrodkowego układu nerwowego organizmu.
Istnieją receptory pierwotne, które są prostymi zakończeniami nerwowymi procesów dośrodkowych komórek nerwowych - neuronów i receptorów wtórnych, które mają wyspecjalizowane komórki do odczuwania pewnego podrażnienia. Receptory pierwotne obejmują na przykład zakończenia nerwowe w skórze, które odbierają bodźce dotykowe i bólowe, a receptory wtórne obejmują komórki węchowe jamy nosowej, czopki i pręciki siatkówki, które odbierają światło. Pręciki to zmodyfikowane komórki nabłonka zawierające substancje, które mogą rozpadać się pod wpływem światła. Powstałe produkty rozpadu powodują zmiany w aktywności tych komórek, które są rejestrowane i przetwarzane przez neurony siatkówki. W zależności od stopnia pobudzenia czopków i pręcików neurony zwiększają lub zmniejszają przepływ impulsów nerwowych wysyłanych do mózgu. Na podobnej zasadzie działają inne wtórne receptory, które odbierają wibracje dźwiękowe, nacisk na skórę i pozycję ciała w przestrzeni.
Istnieją ekstrareceptory (eksteroceptory), które odbierają bodźce zewnętrzne: temperaturę, dotyk, światło, dźwięki, smak, zapach itp .; intrareceptory (interoceptory), które rejestrują stan środowiska wewnętrznego organizmu: skład chemiczny krwi, jej nacisk na ściany naczynia, pracę narządów wewnętrznych; proprioreceptory (proprioceptory), które postrzegają napięcie ścięgien, zmiany długości włókien mięśniowych, aparat więzadłowy. Receptory odbierające wpływy mechaniczne nazywane są mechanoreceptorami, podrażnienia chemiczne chemoreceptorami, a ciśnienie baroreceptorami.
W drugim znaczeniu tego terminu receptory nazywane są odcinkami błony komórkowej, które są wrażliwe na określone substancje i przekazują informację o takim sygnale do komórki. W rzeczywistości receptory błonowe to specjalne cząsteczki białka zdolne do rozpoznawania cząsteczek niektórych związków - białek, peptydów, hormonów o niskiej masie cząsteczkowej, czynników wzrostu i innych substancji. W większości przypadków połączenie receptora z cząsteczką sygnałową aktywuje specjalny enzym. Receptory są rozmieszczone w taki sposób, że cząsteczki, które rozpoznają, lub części tych cząsteczek, mogą wejść do receptorów, jak klucz w dziurce od klucza. Jednocześnie zmienia się stan i aktywność komórki. Na przykład receptory włókien mięśniowych, które zapewniają automatyczną czynność serca, są wrażliwe na hormony - adrenalinę i acetylocholinę. Pierwszy hormon wzmaga czynność serca, drugi ją spowalnia.
Receptory błonowe działają również na połączeniach dwóch komórek nerwowych - synapsach. Zakończenie nerwowe jednej komórki uwalnia specjalną substancję - mediator (na przykład acetylocholinę). Receptory na powierzchni innej komórki odbierają ten sygnał i pobudzają drugą komórkę.
Receptory skórne odpowiadają za naszą zdolność odczuwania dotyku, ciepła, zimna i bólu. Receptory to zmodyfikowane zakończenia nerwowe, które mogą być wolnymi, niewyspecjalizowanymi lub zamkniętymi złożonymi strukturami, które są odpowiedzialne za określony rodzaj wrażliwości. Receptory pełnią rolę sygnalizacyjną, więc są niezbędne, aby człowiek mógł skutecznie i bezpiecznie wchodzić w interakcje ze środowiskiem zewnętrznym.
Główne typy receptorów skórnych i ich funkcje
Wszystkie rodzaje receptorów można podzielić na trzy grupy. Pierwsza grupa receptorów odpowiada za wrażliwość dotykową. Należą do nich ciała Paciniego, Meissnera, Merkel i Ruffiniego. Druga grupa to
termoreceptory: kolby Krausego i wolne zakończenia nerwowe. Trzecia grupa obejmuje receptory bólu.
Dłonie i palce są bardziej wrażliwe na wibracje: ze względu na dużą liczbę receptorów Paciniego w tych obszarach.
Wszystkie typy receptorów mają różne strefy pod względem szerokości czułości, w zależności od pełnionej funkcji.
Receptory skóry:
. receptory skórne odpowiedzialne za wrażliwość dotykową;
. receptory skórne reagujące na zmiany temperatury;
. nocyceptory: receptory skórne odpowiedzialne za wrażliwość na ból.

Receptory skórne odpowiedzialne za wrażliwość na dotyk
Istnieje kilka typów receptorów odpowiedzialnych za wrażenia dotykowe:
. Ciałka Paciniego to receptory, które szybko dostosowują się do zmian ciśnienia i mają szerokie pola receptywne. Receptory te znajdują się w tłuszczu podskórnym i są odpowiedzialne za dużą wrażliwość;
. Ciała Meissnera znajdują się w skórze właściwej i mają wąskie pola odbiorcze, co determinuje ich postrzeganie delikatnej wrażliwości;
. Ciała Merkla - powoli się dostosowują i mają wąskie pola receptorowe, dlatego ich główną funkcją jest wyczuwanie struktury powierzchni;
. Ciała Ruffiniego są odpowiedzialne za uczucie stałego nacisku i znajdują się głównie w okolicy podeszew stóp.
Oddzielnie izolowane są również receptory znajdujące się wewnątrz mieszka włosowego, które sygnalizują odchylenie włosa od jego pierwotnej pozycji.
Receptory skórne reagujące na zmiany temperatury
Według niektórych teorii istnieją różne typy receptorów odczuwania ciepła i zimna. Kolby Krausego odpowiadają za odczuwanie zimna, a wolne zakończenia nerwowe za odczuwanie gorąca. Inne teorie termorecepcji twierdzą, że to wolne zakończenia nerwowe są zaprojektowane do wyczuwania temperatury. W tym przypadku bodźce termiczne są analizowane przez głębokie włókna nerwowe, podczas gdy bodźce zimne są analizowane przez powierzchowne. Receptory wrażliwości na temperaturę tworzą między sobą „mozaikę” składającą się z zimnych i gorących punktów.
Nocyceptory: receptory skórne odpowiedzialne za wrażliwość na ból
Na tym etapie nie ma ostatecznej opinii na temat obecności lub braku receptorów bólu. Niektóre teorie opierają się na fakcie, że za odczuwanie bólu odpowiedzialne są wolne zakończenia nerwowe, które znajdują się w skórze.
Długotrwała i silna stymulacja bólowa stymuluje pojawienie się strumienia impulsów wychodzących, przez co adaptacja do bólu ulega spowolnieniu.
Inne teorie zaprzeczają obecności oddzielnych nocyceptorów. Przyjmuje się, że receptory dotyku i temperatury mają określony próg podrażnienia, powyżej którego pojawia się ból.
Receptory to specyficzne formacje nerwowe, które są zakończeniami wrażliwych (aferentnych) włókien nerwowych, które mogą zostać pobudzone przez działanie bodźca. Receptory odbierające bodźce ze środowiska zewnętrznego nazywane są eksteroceptorami; odbieranie podrażnień ze środowiska wewnętrznego organizmu – interoceptory. Wyróżnia się grupę receptorów zlokalizowanych w mięśniach i ścięgnach szkieletowych oraz sygnalizujących napięcie mięśniowe – proprioreceptory.
W zależności od charakteru bodźca, receptory dzielą się na kilka grup.
1. Mechanoreceptory, które obejmują receptory dotykowe; baroreceptory zlokalizowane w ścianach naczyń krwionośnych i reagujące na zmiany ciśnienia krwi; fonoreceptory reagujące na wibracje powietrza wywołane bodźcem dźwiękowym; receptory aparatu otolitycznego, dostrzegające zmiany położenia ciała w przestrzeni.
2. Chemoreceptory reagujące na działanie jakichkolwiek chemikaliów. Należą do nich osmoreceptory i glukoreceptory, które dostrzegają odpowiednio zmiany ciśnienia osmotycznego i poziomu cukru we krwi; receptory smaku i węchu, które wyczuwają obecność substancji chemicznych w środowisku.
3. Termoreceptory odbierające zmiany temperatury zarówno wewnątrz ciała, jak iw otaczającym go środowisku.
4. Fotoreceptory znajdujące się w siatkówce oka odbierają bodźce świetlne.
5. Receptory bólu wyróżniają się w specjalnej grupie. Mogą być wzbudzane przez mechaniczne, chemiczne i termiczne czynniki drażniące o takiej sile, że możliwe jest ich niszczące działanie na tkanki lub narządy.
Morfologicznie receptory mogą mieć postać prostych wolnych zakończeń nerwowych lub mieć postać włosków, spiral, płytek, podkładek, kulek, stożków, pręcików. Budowa receptorów jest ściśle związana ze specyficznością adekwatnych bodźców, na które receptory mają wysoką bezwzględną czułość. Do pobudzenia fotoreceptorów wystarczy zaledwie 5-10 kwantów światła, a do pobudzenia receptorów węchowych wystarczy jedna cząsteczka substancji zapachowej. Przy przedłużonej ekspozycji na czynnik drażniący dochodzi do adaptacji receptorów, co objawia się zmniejszeniem ich wrażliwości na adekwatny bodziec. Istnieją szybko adaptujące się (dotykowe, baroreceptory) i wolno adaptujące się receptory (chemoreceptory, fonoreceptory). Z kolei przedsionkowo- i proprioreceptory nie dostosowują się. W receptorach pod wpływem bodźca zewnętrznego dochodzi do depolaryzacji błony powierzchniowej, co określa się jako potencjał receptorowy lub generatorowy. Po osiągnięciu wartości krytycznej powoduje wyładowanie aferentnych impulsów pobudzających we włóknie nerwowym wychodzącym z receptora. Informacje odbierane przez receptory z wewnętrznego i zewnętrznego środowiska ciała są przekazywane wzdłuż doprowadzających dróg nerwowych do ośrodkowego układu nerwowego, gdzie są analizowane (patrz Analizatory).