Stosowana biologia molekularna. Biolog molekularny. Opis stanowiska Metody biologii molekularnej
Postępy w badaniach nad kwasami nukleinowymi i biosyntezą białek doprowadziły do powstania szeregu metod o ogromnym znaczeniu praktycznym w medycynie, rolnictwie i wielu innych gałęziach przemysłu.
Po zbadaniu kodu genetycznego oraz podstawowych zasad przechowywania i wdrażania informacji dziedzicznej rozwój biologii molekularnej zatrzymał się, gdyż zabrakło metod pozwalających manipulować genami, izolować je i zmieniać. Pojawienie się tych metod nastąpiło w latach 1970-1980. Dało to potężny impuls do rozwoju tej dziedziny nauki, która kwitnie do dziś. Metody te dotyczą przede wszystkim pozyskiwania pojedynczych genów i ich wprowadzania do komórek innych organizmów (klonowanie i transgeneza molekularna, PCR), a także metod określania sekwencji nukleotydowej w genach (sekwencjonowanie DNA i RNA). Metody te zostaną omówione bardziej szczegółowo poniżej. Zaczniemy od najprostszej metody podstawowej, elektroforezy, a następnie przejdziemy do metod bardziej złożonych.
ELEKTROFOREZA DNA
Jest to podstawowa metoda pracy z DNA, stosowana wraz z niemal wszystkimi innymi metodami w celu izolacji pożądanych cząsteczek i analizy wyników. Elektroforeza żelowa służy do rozdzielania fragmentów DNA według długości. DNA jest kwasem, w jego cząsteczkach znajdują się reszty kwasu fosforowego, które odszczepiają proton i uzyskują ładunek ujemny (ryc. 1).
Dlatego w pole elektryczne Cząsteczki DNA przemieszczają się w stronę anody – dodatnio naładowanej elektrody. Dzieje się tak w roztworze elektrolitu zawierającym jony nośnika ładunku, dzięki czemu roztwór ten przewodzi prąd. Do rozdzielenia fragmentów stosuje się gęsty żel wykonany z polimerów (agarozy lub poliakryloamidu). Cząsteczki DNA „wplątują się” w nie im bardziej, im są dłuższe, dlatego najdłuższe cząsteczki poruszają się najwolniej, a najkrótsze – najszybciej (ryc. 2). Przed lub po elektroforezie żel poddaje się działaniu barwników wiążących się z DNA i fluoryzujących w świetle ultrafioletowym, uzyskując w żelu wzór pasm (patrz ryc. 3). Aby określić długości fragmentów DNA w próbce, porównuje się je z markerem, czyli zbiorem fragmentów o standardowych długościach osadzonych równolegle na tym samym żelu (ryc. 4).



Najważniejszymi narzędziami pracy z DNA są enzymy dokonujące transformacji DNA w żywych komórkach: polimerazy DNA, ligazy DNA i endonukleazy restrykcyjne, czyli enzymy restrykcyjne. Polimeraza DNA Przeprowadzana jest synteza matrycy DNA, która umożliwia namnażanie DNA w probówce. Ligazy DNA zszyć cząsteczki DNA lub załatać w nich luki. Endonukleazy restrykcyjne, Lub restryktazy, tnie cząsteczki DNA według ściśle określonych sekwencji, co pozwala na wycięcie pojedynczych fragmentów z całkowitej masy DNA. Fragmenty te mogą w niektórych przypadkach zawierać pojedyncze geny.
restryktazy
Sekwencje rozpoznawane przez enzymy restrykcyjne są symetryczne, a przerwy mogą wystąpić w środku takiej sekwencji lub z przesunięciem (w tym samym miejscu w obu niciach DNA). Schemat działania różne rodzaje restryktazę pokazano na ryc. 1. W pierwszym przypadku uzyskuje się końcówki tzw. „tępe”, a w drugim końcówki „lepkie”. W przypadku „lepkich” końcówek dna, łańcuszek jest krótszy od drugiego, powstaje odcinek jednoniciowy o symetrycznej sekwencji, która jest taka sama na obu utworzonych końcach.
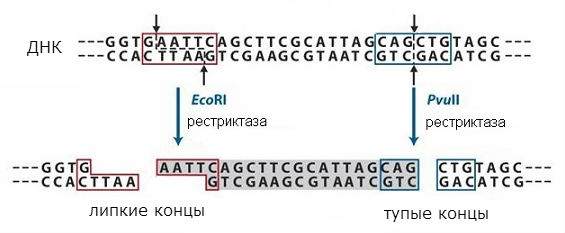
Sekwencje końcowe będą takie same, gdy dowolny DNA zostanie przecięty danym enzymem restrykcyjnym i można je ponownie połączyć, ponieważ mają sekwencje komplementarne. Można je zligować z ligazą DNA, tworząc pojedynczą cząsteczkę. W ten sposób możliwe jest połączenie fragmentów dwóch różnych DNA i uzyskanie tzw rekombinowanego DNA. Podejście to wykorzystywane jest w metodzie klonowania molekularnego, która umożliwia pozyskanie poszczególnych genów i wprowadzenie ich do komórek, które mogą wytworzyć zakodowane w genie białko.
klonowanie molekularne
Klonowanie molekularne wykorzystuje dwie cząsteczki DNA – wstawkę zawierającą gen będący przedmiotem zainteresowania oraz wektor- DNA pełniący rolę nośnika. Wstawkę „wszywuje się” w wektor za pomocą enzymów, uzyskując nową, zrekombinowaną cząsteczkę DNA, następnie cząsteczkę tę wprowadza się do komórek gospodarza, a komórki te tworzą kolonie na pożywce. Kolonia to potomstwo jednej komórki, czyli klon, wszystkie komórki kolonii są genetycznie identyczne i zawierają ten sam rekombinowany DNA. Stąd określenie „klonowanie molekularne”, czyli uzyskanie klonu komórek zawierających interesujący nas fragment DNA. Po uzyskaniu kolonii zawierających interesującą nas wstawkę możemy to zrobić różne metody scharakteryzować tę insercję, na przykład w celu ustalenia jej dokładnej kolejności. Komórki mogą również wytwarzać białko kodowane przez wstawkę, jeśli zawiera ona funkcjonalny gen.
Kiedy do komórek wprowadza się rekombinowaną cząsteczkę, następuje transformacja genetyczna tych komórek. Transformacja- proces wchłaniania przez komórkę organizmu wolnej cząsteczki DNA ze środowiska i jej integracji z genomem, co prowadzi do pojawienia się w takiej komórce nowych dla niej cech dziedzicznych, charakterystycznych dla organizmu-dawcy DNA . Na przykład, jeśli wstawiona cząsteczka zawiera gen oporności na antybiotyk ampicylinę, wówczas transformowane bakterie będą rosły w jego obecności. Przed transformacją ampicylina spowodowała ich śmierć, czyli w transformowanych komórkach pojawił się nowy znak.
WEKTORY
Wektor musi mieć szereg właściwości:
Po pierwsze, jest to stosunkowo mała cząsteczka DNA, którą można łatwo manipulować.
Po drugie, aby DNA mogło zostać zachowane i reprodukowane w komórce, musi zawierać pewną sekwencję zapewniającą jego replikację (początek replikacji lub początek replikacji).
Po trzecie, musi zawierać gen markerowy, co zapewnia selekcję tylko tych komórek, do których wszedł wektor. Zwykle są to geny oporności na antybiotyki – wówczas w obecności antybiotyku wszystkie komórki, które nie zawierają wektora umierają.
Klonowanie genów najczęściej przeprowadza się w komórkach bakteryjnych, ponieważ są one łatwe w hodowli i szybko się namnażają. W komórce bakteryjnej znajduje się zwykle jedna duża, okrągła cząsteczka DNA o długości kilku milionów par zasad, zawierająca wszystkie geny niezbędne bakteriom – chromosom bakteryjny. Oprócz tego u niektórych bakterii występuje małe (kilka tysięcy par zasad) koliste DNA, tzw plazmidy(ryc. 2). Zawierają one, podobnie jak główny DNA, sekwencję nukleotydów, która zapewnia DNA zdolność do replikacji (ori). Plazmidy replikują się niezależnie od głównego (chromosomalnego) DNA, dlatego występują w komórce w dużej liczbie kopii. Wiele z tych plazmidów zawiera geny oporności na antybiotyki, co umożliwia odróżnienie komórek niosących plazmid od komórek normalnych. Częściej stosuje się plazmidy niosące dwa geny nadające oporność na dwa antybiotyki, takie jak tetracyklina i amycylina. Istnieją proste metody izolowania takiego plazmidowego DNA wolnego od DNA głównego chromosomu bakterii.

ZNACZENIE TRANSGENEZY
Nazywa się to przeniesieniem genów z jednego organizmu do drugiego transgeneza, i taki zmodyfikowane organizmy - transgeniczny. Metoda transferu genów do komórek drobnoustrojów służy do otrzymywania rekombinowanych preparatów białkowych do celów leczniczych, w szczególności białek ludzkich, które nie powodują odrzucenia układu odpornościowego – interferonów, insuliny i innych hormonów białkowych, czynników wzrostu komórek, a także białek do produkcji szczepionki. W bardziej skomplikowanych przypadkach, gdy modyfikację białek przeprowadza się prawidłowo tylko w komórkach eukariotycznych, wykorzystuje się hodowle komórek transgenicznych lub zwierzęta transgeniczne, w szczególności zwierzęta gospodarskie (głównie kozy), które wydzielają niezbędne białka do mleka lub białka izoluje się z ich krwi . W ten sposób powstają przeciwciała, czynniki krzepnięcia krwi i inne białka. Metodą transgenezy uzyskuje się rośliny uprawne, które są odporne na herbicydy i szkodniki oraz posiadają inne przydatne właściwości. Wykorzystując mikroorganizmy transgeniczne do oczyszczania ścieków i zwalczania zanieczyszczeń, istnieją nawet drobnoustroje transgeniczne, które mogą rozkładać ropę naftową. Ponadto technologie transgeniczne są niezbędne w badaniach naukowych – rozwój biologii jest dziś nie do pomyślenia bez rutynowego stosowania metod modyfikacji i transferu genów.
technologii klonowania molekularnego
wstawki
Aby uzyskać indywidualny gen z dowolnego organizmu, izoluje się z niego cały chromosomalny DNA i rozszczepia jednym lub dwoma enzymami restrykcyjnymi. Enzymy dobiera się tak, aby nie przecinały interesującego nas genu, lecz robiły przerwy wzdłuż jego krawędzi, a w plazmidowym DNA dokonały jednej przerwy w jednym z genów oporności, np. na ampicylinę.
Proces klonowania molekularnego obejmuje następujące etapy:
Cięcie i zszywanie – konstrukcja pojedynczej rekombinowanej cząsteczki z wstawki i wektora.
Transformacja polega na wprowadzeniu rekombinowanej cząsteczki do komórek.
Selekcja - selekcja komórek, które otrzymały wektor z wstawką.
cięcie i szycie
Plazmidowy DNA jest traktowany tymi samymi enzymami restrykcyjnymi i przekształca się w cząsteczkę liniową, jeśli zostanie wybrany taki enzym restrykcyjny, który wprowadza 1 przerwę do plazmidu. W rezultacie te same lepkie końce pojawiają się na końcach wszystkich powstałych fragmentów DNA. W miarę obniżania temperatury końce te łączą się losowo i ligazą DNA (patrz ryc. 3).

Otrzymuje się mieszaninę kolistych DNA o różnym składzie: niektóre z nich będą zawierały określoną sekwencję DNA chromosomalnego DNA połączoną z DNA bakteryjnym, inne będą zawierać połączone ze sobą fragmenty chromosomalnego DNA, a jeszcze inne będą zawierać zredukowany plazmid kolisty lub jego dimer (ryc. 4).

transformacja
Następnie przeprowadza się tę mieszaninę transformacja genetyczna bakterie nie zawierające plazmidów. Transformacja- proces wchłaniania przez komórkę organizmu wolnej cząsteczki DNA ze środowiska i jej integracji z genomem, co prowadzi do pojawienia się w takiej komórce nowych dla niej cech dziedzicznych, charakterystycznych dla organizmu-dawcy DNA . Tylko jeden plazmid może wejść i rozmnażać się w każdej komórce. Takie komórki umieszcza się na stałej pożywce zawierającej antybiotyk tetracyklinę. Komórki, które nie otrzymały plazmidu, nie będą rosły na tym podłożu, a komórki niosące plazmid tworzą kolonie, z których każda zawiera potomków tylko jednej komórki, tj. wszystkie komórki w kolonii niosą ten sam plazmid (patrz ryc. 5).

Wybór
Następnie zadaniem jest wyizolowanie tylko tych komórek, do których wszedł wektor z wstawką i odróżnienie ich od komórek niosących tylko wektor bez wstawki lub nieniosących wektora w ogóle. Ten proces wybierania właściwych komórek nazywa się wybór. W tym celu zastosuj markery selektywne- zwykle geny oporności na antybiotyki w wektorze, oraz media selektywne zawierające antybiotyki lub inne substancje selektywne.
W rozważanym przez nas przykładzie komórki z kolonii hodowanych w obecności ampicyliny hoduje się na dwóch podłożach: pierwsze zawiera ampicylinę, a drugie zawiera tetracyklinę. Kolonie zawierające tylko plazmid będą rosły na obu podłożach, natomiast kolonie zawierające wstawiony chromosomalny DNA do plazmidów nie będą rosły na podłożu z tetracykliną (ryc. 5). Spośród nich te, które zawierają interesujący nas gen, wybiera się specjalnymi metodami, hoduje w wystarczających ilościach i izoluje plazmidowy DNA. Z niego, przy użyciu tych samych restryktaz, które zastosowano do uzyskania rekombinowanego DNA, wycina się indywidualny gen będący przedmiotem zainteresowania. DNA tego genu można wykorzystać do ustalenia sekwencji nukleotydów, wprowadzić do dowolnego organizmu w celu uzyskania nowych właściwości lub zsyntetyzować pożądane białko. Ta metoda izolacji genów nazywa się klonowanie molekularne.
FLUORESCENCYJNE BIAŁKA
Bardzo wygodne jest stosowanie białek fluorescencyjnych jako genów markerowych w badaniach organizmów eukariotycznych. Gen pierwszego białka fluorescencyjnego, białko zielonej fluorescencji (GFP) został wyizolowany z meduzy Aqeuorea victoria i wprowadzony do różnych organizmów modelowych (patrz ryc. 6). W 2008 roku O. Shimomura, M. Chalfi i R. Tsien otrzymali Nagrodę Nobla za odkrycie i zastosowanie tego białka.

Następnie wyizolowano geny innych białek fluorescencyjnych – czerwonego, niebieskiego, żółtego. Geny te zostały sztucznie zmodyfikowane w celu wytwarzania białek pożądane właściwości. Różnorodność białek fluorescencyjnych pokazano na ryc. 7, która przedstawia szalkę Petriego z bakteriami zawierającymi geny różnych białek fluorescencyjnych.

zastosowanie białek fluorescencyjnych
Gen białka fluorescencyjnego można poddać fuzji z genem dowolnego innego białka, wtedy podczas translacji powstanie pojedyncze białko – translacyjne białko fuzyjne, lub połączenie(białko fuzyjne), które fluoryzuje. W ten sposób można na przykład badać lokalizację (lokalizację) dowolnych białek będących przedmiotem zainteresowania w komórce, ich ruch. Stosując ekspresję białek fluorescencyjnych tylko w niektórych typach komórek, możliwe jest oznaczenie komórek tego typu w organizmie wielokomórkowym (patrz ryc. 8 - mózg myszy, w którym poszczególne neurony mają różne kolory ze względu na pewną kombinację genów białek fluorescencyjnych). Białka fluorescencyjne są niezbędnym narzędziem współczesnej biologii molekularnej.

PCR
Inną metodą pozyskiwania genów jest tzw reakcja łańcuchowa polimerazy (PCR). Opiera się na zdolności polimeraz DNA do uzupełnienia drugiej nici DNA wzdłuż nici komplementarnej, tak jak to ma miejsce w komórkach podczas replikacji DNA.
Początki replikacji w tej metodzie są wyznaczane przez dwa małe fragmenty DNA zwane posiew, Lub podkłady. Startery te są komplementarne do końców interesującego genu na dwóch niciach DNA. Najpierw chromosomalny DNA, z którego ma zostać wyizolowany gen, miesza się z nasionami i podgrzewa do temperatury 99°C. Prowadzi to do zerwania wiązań wodorowych i rozbieżności nici DNA. Następnie temperaturę obniża się do 50-70 około C (w zależności od długości i kolejności nasion). W tych warunkach startery przyłączają się do komplementarnych regionów chromosomalnego DNA, tworząc regularną podwójną helisę (patrz ryc. 9). Następnie dodaje się mieszaninę wszystkich czterech nukleotydów potrzebnych do syntezy DNA i polimerazy DNA. Enzym wydłuża startery budując dwuniciowy DNA od miejsca przyłączenia starterów, tj. od końców genu do końca jednoniciowej cząsteczki chromosomu. 
Jeśli mieszaninę ponownie podgrzeje się, łańcuchy chromosomalne i nowo zsyntetyzowane ulegną rozproszeniu. Po ostygnięciu ponownie dołączą się do nich nasiona, które są pobierane w dużym nadmiarze (patrz ryc. 10).

Na nowo syntetyzowanych łańcuchach połączą się one nie z końcem, od którego rozpoczęła się pierwsza synteza, ale z przeciwległym, ponieważ łańcuchy DNA są antyrównoległe. Dlatego w drugim cyklu syntezy na takich łańcuchach zostanie uzupełniona jedynie sekwencja odpowiadająca genowi (patrz ryc. 11).

W metodzie tej wykorzystuje się polimerazę DNA z bakterii termofilnych, która wytrzymuje gotowanie i działa w temperaturze 70-80°C, nie trzeba jej dodawać za każdym razem, ale wystarczy dodać ją na początku doświadczenia. Powtarzając procedury ogrzewania i chłodzenia w tej samej kolejności, możemy podwoić liczbę sekwencji w każdym cyklu, ograniczoną na obu końcach wprowadzonymi nasionami (patrz ryc. 12).

Po około 25 takich cyklach liczba kopii genu wzrośnie ponad milion razy. Takie ilości można łatwo oddzielić od wprowadzonego do probówki chromosomalnego DNA i wykorzystać do różnych celów.
sekwencjonowanie DNA
Kolejnym ważnym osiągnięciem jest opracowanie metod określania sekwencji nukleotydów w DNA - sekwencjonowanie DNA(z angielskiej sekwencji - sekwencja). W tym celu konieczne jest uzyskanie genów czystych z innego DNA jedną z opisanych metod. Następnie łańcuchy DNA rozdziela się poprzez ogrzewanie i dodaje do nich starter znakowany radioaktywnym fosforem lub znacznikiem fluorescencyjnym. Należy pamiętać, że pobierane jest jedno ziarno, uzupełniające jeden łańcuch. Następnie dodaje się polimerazę DNA i mieszaninę 4 nukleotydów. Taką mieszaninę dzieli się na 4 części i do każdej dodaje się jeden z nukleotydów, zmodyfikowany tak, aby nie zawierał grupy hydroksylowej na trzecim atomie dezoksyrybozy. Jeśli taki nukleotyd zostanie włączony do syntetyzowanego łańcucha DNA, wówczas jego wydłużanie nie będzie mogło być kontynuowane, ponieważ polimeraza nie będzie miała gdzie przyłączyć kolejnego nukleotydu. Dlatego synteza DNA po włączeniu takiego nukleotydu zostaje przerwana. Nukleotydy te, zwane dideoksynukleotydami, dodaje się znacznie rzadziej niż zwykle, dlatego zakończenie łańcucha następuje tylko sporadycznie i w każdym łańcuchu w różnych miejscach. Rezultatem jest mieszanina łańcuchów różne długości, na końcu każdego z nich znajduje się ten sam nukleotyd. Zatem długość łańcucha odpowiada liczbie nukleotydów w badanej sekwencji, np. jeśli mieliśmy dideoksynukleotyd adenylowy, a powstałe łańcuchy miały długość 2, 7 i 12 nukleotydów, to adenina znajdowała się na drugiej, siódmej i dwunastej pozycji gen. Powstałą mieszaninę łańcuchów można łatwo rozdzielić według wielkości za pomocą elektroforezy, a zsyntetyzowane łańcuchy można zidentyfikować za pomocą radioaktywności na kliszy rentgenowskiej (patrz ryc. 10).

Okazuje się, że zdjęcie pokazane na dole zdjęcia nazywa się radioautografem. Poruszając się po nim od dołu do góry i czytając literę nad kolumnami każdej strefy, otrzymamy sekwencję nukleotydów pokazaną na rysunku po prawej stronie autografu. Okazało się, że syntezę zatrzymują nie tylko dideoksynukleotydy, ale także nukleotydy, w których na trzeciej pozycji cukru jest przyłączona jakaś grupa chemiczna, np. barwnik fluorescencyjny. Jeśli każdy nukleotyd zostanie oznakowany własnym barwnikiem, wówczas strefy powstałe w wyniku rozdzielenia syntetyzowanych łańcuchów będą świecić innym światłem. Umożliwia to prowadzenie reakcji w jednej probówce jednocześnie dla wszystkich nukleotydów i rozdzielanie powstałych łańcuchów według długości, identyfikację nukleotydów według koloru (patrz ryc. 11).

Metody te umożliwiły określenie sekwencji nie tylko poszczególnych genów, ale także odczytanie całych genomów. Obecnie opracowano jeszcze szybsze metody określania sekwencji nukleotydów w genach. Jeśli pierwszy ludzki genom został rozszyfrowany przez duże międzynarodowe konsorcjum przy użyciu pierwszej podanej metody w ciągu 12 lat, drugi przy użyciu drugiej w ciągu trzech lat, teraz można to zrobić w ciągu miesiąca. Pozwala to przewidzieć predyspozycje danej osoby do wielu chorób i z wyprzedzeniem podjąć działania, aby ich uniknąć.
Komiks na konkurs „bio/mol/tekst”: Dziś probówka biologa molekularnego poprowadzi Cię przez świat niesamowitej nauki - biologii molekularnej! Zaczniemy od historycznej wycieczki po etapach jego rozwoju, opiszemy główne odkrycia i eksperymenty od 1933 roku. Opiszemy także jasno główne metody biologii molekularnej, które umożliwiły manipulowanie genami, zmianę ich i izolację. Pojawienie się tych metod stanowiło silny impuls do rozwoju biologii molekularnej. Pamiętajmy też o roli biotechnologii i poruszajmy jeden z najpopularniejszych tematów w tym obszarze – edycję genomu z wykorzystaniem systemów CRISPR/Cas.
Sponsorem generalnym konkursu i partnerem nominacji Skoltech jest firma .
Sponsorem konkursu jest firma Diaem: największy dostawca sprzętu, odczynników i materiałów eksploatacyjnych do badań i produkcji biologicznej.

Firma była sponsorem nagrody publiczności.
„Książka” sponsorem konkursu – „Alpina non-fiction”
1. Wstęp. Istota biologii molekularnej

Zajmuje się badaniem podstaw czynności życiowych organizmów na poziomie makrocząsteczek. Celem biologii molekularnej jest ustalenie roli i mechanizmów funkcjonowania tych makrocząsteczek w oparciu o wiedzę o ich budowie i właściwościach.
Historycznie rzecz biorąc, biologia molekularna powstała podczas rozwoju dziedzin biochemii zajmujących się badaniem kwasów nukleinowych i białek. Chociaż biochemia zajmuje się badaniem metabolizmu, skład chemicznyżywe komórki, organizmy i zachodzące w nich procesy chemiczne, biologia molekularna skupia się na badaniu mechanizmów przekazywania, reprodukcji i przechowywania informacji genetycznej.
Przedmiotem badań biologii molekularnej są same kwasy nukleinowe - dezoksyrybonukleinowy (DNA), rybonukleinowy (RNA) - i białka, a także ich kompleksy makromolekularne - chromosomy, rybosomy, układy wieloenzymowe, które zapewniają biosyntezę białek i kwasów nukleinowych. Biologia molekularna graniczy również z przedmiotami badań i częściowo pokrywa się z genetyką molekularną, wirusologią, biochemią i szeregiem innych pokrewnych nauk biologicznych.
2. Wycieczka historyczna przez etapy rozwoju biologii molekularnej

Jako odrębna dziedzina biochemii biologia molekularna zaczęła się rozwijać w latach 30. ubiegłego wieku. Już wtedy konieczne stało się zrozumienie fenomenu życia na poziomie molekularnym, aby zbadać procesy przekazywania i przechowywania informacji genetycznej. Właśnie w tym czasie postawiono zadanie biologii molekularnej w badaniu właściwości, struktury i interakcji białek i kwasów nukleinowych.
Termin „biologia molekularna” został po raz pierwszy użyty w r 1933 rok William Astbury podczas badań białek fibrylarnych (kolagen, fibryna krwi, białka mięśni kurczliwych). Astbury badał związek między strukturą molekularną a biologicznymi, fizycznymi właściwościami tych białek. Na początku powstania biologii molekularnej uważano, że RNA jest składnikiem wyłącznie roślin i grzybów, a DNA – wyłącznie zwierząt. I w 1935 Odkrycie DNA grochu przez Andrieja Biełozerskiego doprowadziło do ustalenia faktu, że DNA znajduje się w każdej żywej komórce.
W 1940 Kolosalnym osiągnięciem było ustalenie przez George'a Beadle'a i Edwarda Tathama związku przyczynowego między genami i białkami. Hipoteza naukowców „Jeden gen – jeden enzym” stała się podstawą koncepcji, że specyficzna struktura białka jest regulowana przez geny. Uważa się, że informacja genetyczna jest kodowana przez specjalną sekwencję nukleotydów w DNA, która reguluje pierwotną strukturę białek. Później udowodniono, że wiele białek ma strukturę czwartorzędową. W tworzeniu takich struktur biorą udział różne łańcuchy peptydowe. Na tej podstawie przepis dotyczący związku między genem a enzymem został nieco przekształcony i obecnie brzmi jak „Jeden gen – jeden polipeptyd”.
W 1944 W 1999 roku amerykański biolog Oswald Avery i jego współpracownicy (Colin McLeod i McLean McCarthy) udowodnili, że substancją powodującą transformację bakterii jest DNA, a nie białka. Eksperyment posłużył jako dowód na rolę DNA w przekazywaniu informacji dziedzicznej, przekreślając przestarzałą wiedzę na temat białkowej natury genów.

We wczesnych latach pięćdziesiątych Frederick Sanger wykazał, że łańcuch białkowy to unikalna sekwencja reszt aminokwasowych. W 1951 I 1952 lat naukowiec ustalił pełną sekwencję dwóch łańcuchów polipeptydowych – insuliny bydlęcej W(30 reszt aminokwasowych) i A(odpowiednio 21 reszt aminokwasowych).
Mniej więcej w tym samym czasie, w 1951–1953 Erwin Chargaff sformułował zasady dotyczące proporcji zasad azotowych w DNA. Zgodnie z zasadą, niezależnie od różnic gatunkowych organizmów żywych w ich DNA, ilość adeniny (A) jest równa ilości tyminy (T), a ilość guaniny (G) jest równa ilości cytozyny (C).
W 1953 udowodnił genetyczną rolę DNA. James Watson i Francis Crick na podstawie zdjęcia rentgenowskiego DNA uzyskanego przez Rosalind Franklin i Maurice’a Wilkinsa ustalili strukturę przestrzenną DNA i wysunęli potwierdzone później założenie dotyczące mechanizmu jego replikacji (podwojenia), który leży u podstaw dziedziczności.

1958 rok - powstanie przez Francisa Cricka centralnego dogmatu biologii molekularnej: transfer informacji genetycznej przebiega w kierunku DNA → RNA → białko.
Istota dogmatu polega na tym, że w komórkach następuje pewien ukierunkowany przepływ informacji z DNA, które z kolei jest oryginalnym tekstem genetycznym, składającym się z czterech liter: A, T, G i C. Jest on zapisany w DNA podwójna helisa w postaci sekwencji tych liter - nukleotydów.
Ten tekst jest w trakcie transkrypcji. I proces ten nazywa się transkrypcja. Podczas tego procesu syntetyzowany jest RNA, który jest identyczny z tekstem genetycznym, z tą różnicą, że w RNA zamiast T występuje U (uracyl).
To RNA nazywa się informacyjny RNA (mRNA), Lub matryca (mRNA). Audycja mRNA przeprowadza się przy użyciu kodu genetycznego w postaci sekwencji tripletowych nukleotydów. Podczas tego procesu tekst kwasów nukleinowych DNA i RNA jest tłumaczony z czteroliterowego tekstu na dwudziestoliterowy tekst aminokwasów.
Jest tylko dwadzieścia naturalnych aminokwasów, a w tekście kwasów nukleinowych znajdują się cztery litery. Z tego powodu istnieje tłumaczenie z alfabetu czteroliterowego na alfabet dwudziestoliterowy poprzez kod genetyczny, w którym każde trzy nukleotydy odpowiadają aminokwasowi. Można więc z czterech liter ułożyć całe 64 trzyliterowe kombinacje, w dodatku aminokwasów jest 20. Z tego wynika, że kod genetyczny musi koniecznie posiadać właściwość degeneracji. Jednak w tym czasie kod genetyczny nie był znany, poza tym nawet nie zaczęto go rozszyfrowywać, ale Crick sformułował już swój główny dogmat.
Niemniej jednak istniała pewność, że kod musi istnieć. Do tego czasu udowodniono, że kod ten miał znak trójkowy. Oznacza to, że konkretnie trzy litery w kwasach nukleinowych ( kodony) odpowiadają dowolnemu aminokwasowi. Jest ich 64, kodują one 20 aminokwasów. Oznacza to, że każdemu aminokwasowi odpowiada kilka kodonów jednocześnie.
Można zatem stwierdzić, że centralnym dogmatem jest postulat mówiący, że w komórce następuje ukierunkowany przepływ informacji: DNA → RNA → białko. Crick podkreślił główną treść centralnego dogmatu: nie może nastąpić odwrotny przepływ informacji, białko nie jest zdolne do zmiany informacji genetycznej.
Oto główne znaczenie centralnego dogmatu: białko nie jest w stanie zmieniać i przekształcać informacji w DNA (lub RNA), przepływ zawsze odbywa się tylko w jednym kierunku.
Jakiś czas później odkryto nowy enzym, który nie był znany w momencie formułowania centralnego dogmatu: odwrotna transkryptaza który syntetyzuje DNA z RNA. Enzym odkryto w wirusach, w których informacja genetyczna jest zakodowana w RNA, a nie w DNA. Takie wirusy nazywane są retrowirusami. Mają otoczkę wirusową z zamkniętym w niej RNA i specjalny enzym. Enzym jest odwrotną transkryptazą, która syntetyzuje DNA zgodnie z matrycą tego wirusowego RNA, a DNA ten służy następnie jako materiał genetyczny do dalszego rozwoju wirusa w komórce.
Oczywiście to odkrycie wywołało wielki szok i wiele kontrowersji wśród biologów molekularnych, ponieważ uważano, że w oparciu o centralny dogmat tak nie może być. Jednak Crick od razu wyjaśnił, że nigdy nie powiedział, że to niemożliwe. Powiedział tylko, że nigdy nie może nastąpić przepływ informacji z białka do kwasów nukleinowych, a już wewnątrz kwasów nukleinowych możliwe są wszelkiego rodzaju procesy: synteza DNA na DNA, DNA na RNA, RNA na DNA i RNA na RNA.
Po sformułowaniu centralnego dogmatu nadal pozostawało wiele pytań: w jaki sposób alfabet czterech nukleotydów tworzących DNA (lub RNA) koduje 20-literowy alfabet aminokwasów tworzących białka? Jaka jest istota kodu genetycznego?
Pierwsze poglądy na temat istnienia kodu genetycznego sformułował Alexander Downes ( 1952 d.) i Georgy Gamov ( 1954 G.). Naukowcy wykazali, że sekwencja nukleotydów musi zawierać co najmniej trzy połączenia. Później udowodniono, że taka sekwencja składa się z trzech nukleotydów, tzw kodon (tryplet). Jednakże kwestia, które nukleotydy są odpowiedzialne za włączenie którego aminokwasu do cząsteczki białka, pozostawała otwarta aż do 1961 roku.

I w 1961 Marshall Nirenberg wraz z Heinrichem Mattei używali systemu do nadawania in vitro. Jako matrycę zastosowano oligonukleotyd. Zawierał jedynie reszty uracylu, a syntetyzowany z niego peptyd zawierał jedynie aminokwas fenyloalaninę. W ten sposób po raz pierwszy ustalono znaczenie kodonu: kodon UUU koduje fenyloalaninę. Później Har Koran odkrył, że sekwencja nukleotydów UCUCUCUCUCUC koduje zestaw aminokwasów seryna-leucyna-seryna-leucyna. W zasadzie dzięki dziełom Nirenberga i Koranu 1965 roku kod genetyczny został całkowicie rozwikłany. Okazało się, że każdy triplet koduje konkretny aminokwas. Kolejność kodonów określa kolejność aminokwasów w białku.
Główne zasady funkcjonowania białek i kwasów nukleinowych zostały sformułowane na początku lat 70-tych. Stwierdzono, że synteza białek i kwasów nukleinowych przebiega zgodnie z mechanizmem macierzowym. Cząsteczka szablonowa niesie zakodowaną informację o sekwencji aminokwasów lub nukleotydów. Podczas replikacji lub transkrypcji matrycą jest DNA, a podczas translacji i odwrotnej transkrypcji jest to mRNA.
W ten sposób stworzono przesłanki do powstania dziedzin biologii molekularnej, w tym inżynierii genetycznej. W 1972 roku Paul Berg i współpracownicy opracowali technologię klonowania molekularnego. Naukowcy uzyskali pierwszy rekombinowany DNA in vitro. Te wybitne odkrycia stworzyły podstawę nowego kierunku w biologii molekularnej i 1972 od tego czasu rok ten uznano za datę narodzin inżynierii genetycznej.



3. Metody biologii molekularnej

Ogromny postęp w badaniach nad kwasami nukleinowymi, strukturą DNA i biosyntezą białek doprowadził do powstania szeregu metod mających ogromne znaczenie w medycynie, rolnictwie i nauce w ogóle.
Po przestudiowaniu kodu genetycznego oraz podstawowych zasad przechowywania, przekazywania i wdrażania informacji dziedzicznej konieczne stały się specjalne metody dla dalszego rozwoju biologii molekularnej. Metody te umożliwiłyby manipulowanie, modyfikowanie i izolację genów.
Pojawienie się takich metod nastąpiło w latach 70. i 80. XX wieku. Dało to ogromny impuls do rozwoju biologii molekularnej. Przede wszystkim metody te są bezpośrednio związane z wytwarzaniem genów i ich wprowadzaniem do komórek innych organizmów, a także możliwością określenia sekwencji nukleotydów w genach.
3.1. Elektroforeza DNA

Elektroforeza DNA to podstawowa metoda pracy z DNA. Elektroforeza DNA jest stosowana wraz z prawie wszystkimi innymi metodami w celu wyizolowania pożądanych cząsteczek i dalszej analizy wyników. Sama metoda elektroforezy żelowej służy do rozdzielania fragmentów DNA według długości.


Przed lub po elektroforezie żel poddaje się działaniu barwników, które mogą wiązać się z DNA. Barwniki fluoryzują w świetle ultrafioletowym, tworząc wzór pasm w żelu. Aby określić długość fragmentów DNA, można je porównać znaczniki- zestawy fragmentów o standardowych długościach, które nakładamy na ten sam żel.

Fluorescencyjne białka
Podczas badania organizmów eukariotycznych wygodnie jest stosować białka fluorescencyjne jako geny markerowe. Gen pierwszego białka zielonej fluorescencji ( białko zielonej fluorescencji, GFP) wyizolowany z meduz Aqeuorea Wiktoria a następnie wprowadzane do różnych organizmów. Następnie wyizolowano geny białek fluorescencyjnych o innych kolorach: niebieskim, żółtym, czerwonym. Aby uzyskać białka o interesujących właściwościach, takie geny zostały sztucznie zmodyfikowane.

Generalnie najważniejszymi narzędziami pracy z cząsteczką DNA są enzymy dokonujące szeregu transformacji DNA w komórkach: Polimeraza DNA, Ligazy DNA I restryktazy (endonukleazy restrykcyjne).
transgeneza
transgeneza Nazywa się to przeniesieniem genów z jednego organizmu do drugiego. Takie organizmy nazywane są transgeniczny.
Rekombinowane preparaty białkowe uzyskuje się właśnie poprzez przeniesienie genów do komórek mikroorganizmu. Większość tych białek to tzw interferony, insulina, niektóre hormony białkowe, a także białka do produkcji szeregu szczepionek.
W innych przypadkach wykorzystuje się hodowle komórkowe eukariontów lub zwierząt transgenicznych, głównie hodowlanych, które wydzielają do mleka niezbędne białka. W ten sposób uzyskuje się przeciwciała, czynniki krzepnięcia krwi i inne białka. Metodą transgenezy uzyskuje się plony odporne na szkodniki i herbicydy, a ścieki oczyszcza się za pomocą mikroorganizmów transgenicznych.
Oprócz tego, technologie transgeniczne są niezbędne w badaniach naukowych, ponieważ rozwój biologii jest szybszy dzięki zastosowaniu metod modyfikacji i transferu genów.

Ograniczacze
Sekwencje rozpoznawane przez enzymy restrykcyjne są symetryczne, więc wszelkiego rodzaju pęknięcia mogą wystąpić albo w środku takiej sekwencji, albo wraz z przesunięciem jednej lub obu nici cząsteczki DNA.
Podczas dzielenia dowolnego DNA za pomocą enzymu restrykcyjnego sekwencje na końcach fragmentów będą takie same. Będą mogli ponownie się połączyć, ponieważ mają uzupełniające się witryny.
Możesz uzyskać pojedynczą cząsteczkę, łącząc te sekwencje za pomocą Ligazy DNA. Dzięki temu możliwe jest połączenie fragmentów dwóch różnych DNA i otrzymanie rekombinowanego DNA.


3.2. PCR
Metoda opiera się na zdolności polimeraz DNA do kompletowania drugiej nici DNA wzdłuż nici komplementarnej w taki sam sposób, jak w procesie replikacji DNA w komórce.



3.3. sekwencjonowanie DNA
Szybki rozwój metody sekwencjonowania pozwala skutecznie określić cechy badanego organizmu już na poziomie jego genomu. Główną zaletą takich technologii genomowych i postgenomicznych jest zwiększenie możliwości badawczych i studyjnych. charakter genetyczny choroby ludzkie, w celu wstępnego podjęcia niezbędne środki i unikaj chorób.
Dzięki badaniom na dużą skalę możliwe jest uzyskanie niezbędnych danych na temat różnych cech genetycznych różnych grup ludzi, a tym samym rozwój metod medycyny. Z tego powodu identyfikacja genetycznej predyspozycji do różnych chorób jest dziś bardzo popularna.
Podobne metody mają szerokie zastosowanie praktycznie na całym świecie, także w Rosji. W związku z postępem naukowym metody tego typu są wprowadzane do badań medycznych i m.in praktyka lekarska ogólnie.



4. Biotechnologia

Biotechnologia- dyscyplina badająca możliwości wykorzystania organizmów żywych lub ich układów do rozwiązywania problemów technologicznych, a także tworzenia organizmów żywych o pożądanych właściwościach poprzez inżynierię genetyczną. Biotechnologia wykorzystuje metody chemii, mikrobiologii, biochemii i oczywiście biologii molekularnej.
Główne kierunki rozwoju biotechnologii (wprowadzane są zasady procesów biotechnologicznych do produkcji wszystkich gałęzi przemysłu):
- Tworzenie i produkcja nowych rodzajów żywności i pasz dla zwierząt.
- Pozyskiwanie i badanie nowych szczepów mikroorganizmów.
- Hodowla nowych odmian roślin, a także tworzenie środków ochrony roślin przed chorobami i szkodnikami.
- Zastosowanie metod biotechnologicznych dla potrzeb ekologii. Takie metody biotechnologii stosuje się do recyklingu, usuwania odpadów, czyszczenia Ścieki, wywiewu i sanitacji gleb.
- Produkcja witamin, hormonów, enzymów, surowic na potrzeby medycyny. Biotechnolodzy rozwijają się coraz lepiej leki wcześniej uważana za nieuleczalną.
Najważniejszym osiągnięciem biotechnologii jest inżynieria genetyczna.
Inżynieria genetyczna- zespół technologii i metod otrzymywania rekombinowanych cząsteczek RNA i DNA, izolowania poszczególnych genów z komórek, manipulacji genami i wprowadzania ich do innych organizmów (bakterie, drożdże, ssaki). Organizmy takie są w stanie wytworzyć produkty końcowe o pożądanych, zmodyfikowanych właściwościach.
Metody inżynierii genetycznej mają na celu konstruowanie nowych, nieistniejących wcześniej w przyrodzie kombinacji genów.
Mówiąc o osiągnięciach inżynierii genetycznej, nie sposób nie poruszyć tematu klonowania. Klonowanie jest jedną z metod biotechnologii wykorzystywaną do uzyskania identycznego potomstwa różnych organizmów poprzez rozmnażanie bezpłciowe.
Innymi słowy, klonowanie można traktować jako proces tworzenia genetycznie identycznych kopii organizmu lub komórki. A sklonowane organizmy są podobne lub całkowicie identyczne nie tylko pod względem cech zewnętrznych, ale także treści genetycznej.
Osławiona owca Dolly w 1966 roku została pierwszym sklonowanym ssakiem. Uzyskano go poprzez przeszczepienie jądra komórki somatycznej do cytoplazmy komórki jajowej. Dolly była genetyczną kopią owcy-dawcy jądra. W naturalnych warunkach osobnik powstaje z jednego zapłodnionego jaja, otrzymując połowę materiału genetycznego od dwojga rodziców. Jednakże podczas klonowania materiał genetyczny pobrano z komórki jednego osobnika. Najpierw z zygoty usunięto jądro zawierające samo DNA. Następnie usunęli jądro z komórki dorosłej owcy i wszczepili je do zygoty bez jądra, a następnie przeszczepili je do macicy dorosłej osoby i pozwolono im rosnąć i rozwijać się.

Jednak nie wszystkie próby klonowania zakończyły się sukcesem. Równolegle z klonowaniem Dolly przeprowadzono eksperyment polegający na zastąpieniu DNA na 273 innych jajach. Ale tylko w jednym przypadku żywe dorosłe zwierzę mogło w pełni się rozwinąć i rosnąć. Po Dolly naukowcy próbowali sklonować inne typy ssaków.

Jednym z rodzajów inżynierii genetycznej jest edycja genomu.
Narzędzie CRISPR/Cas opiera się na elemencie układu odpornościowego bakterii, który naukowcy zaadaptowali do wprowadzania wszelkich zmian w DNA zwierząt czy roślin.


CRISPR/Cas to jedna z biotechnologicznych metod manipulacji pojedynczymi genami w komórkach. Istnieje wiele zastosowań tej technologii. CRISPR/Cas umożliwia naukowcom poznanie funkcji różnych genów. Aby to zrobić, wystarczy wyciąć badany gen z DNA i zbadać, które funkcje organizmu zostały dotknięte.
Kilka praktycznych zastosowań systemu:
- Rolnictwo. Dzięki systemom CRISPR/Cas można ulepszyć plony. Mianowicie po to, aby były smaczniejsze i pożywniejsze, a także odporne na ciepło. Można nadać roślinom inne właściwości: na przykład wyciąć gen alergenu z orzechów (orzeszków ziemnych lub orzechów laskowych).
- Medycyna, choroby dziedziczne. Celem naukowców jest wykorzystanie CRISPR/Cas do usunięcia mutacji z ludzkiego genomu, które mogą powodować choroby, takie jak anemia sierpowatokrwinkowa itp. Teoretycznie CRISPR/Cas może zatrzymać rozwój wirusa HIV.
- Napęd genowy. CRISPR/Cas może zmienić nie tylko genom pojedynczego zwierzęcia lub rośliny, ale także pulę genową gatunku. Koncepcja ta znana jest jako „napęd genowy”. Każdy żywy organizm przekazuje potomstwu połowę swoich genów. Jednak użycie CRISPR/Cas może zwiększyć szansę na transfer genu nawet o 100%. Jest to ważne, aby pożądana cecha szybciej rozprzestrzeniła się w całej populacji.
Szwajcarscy naukowcy znacząco udoskonalili i unowocześnili metodę edycji genomu CRISPR/Cas, poszerzając tym samym jej możliwości. Jednakże naukowcy mogli modyfikować tylko jeden gen na raz, korzystając z systemu CRISPR/Cas. Ale teraz naukowcy z ETH Zurich opracowali metodę, która może jednocześnie modyfikować 25 genów w komórce.
W przypadku najnowszej techniki eksperci wykorzystali enzym Cas12a. Genetykom po raz pierwszy w historii udało się sklonować małpy. „Popularna mechanika”;
Biologia molekularna przeżyła okres szybkiego rozwoju własnych metod badawczych, które obecnie różnią się od biochemii. Należą do nich w szczególności metody inżynierii genetycznej, klonowania, sztucznej ekspresji i nokautu genów. Ponieważ DNA jest materialnym nośnikiem informacji genetycznej, biologia molekularna znacznie zbliżyła się do genetyki, a na styku powstała genetyka molekularna, która jest jednocześnie działem genetyki i biologii molekularnej. Tak jak biologia molekularna szeroko wykorzystuje wirusy jako narzędzie badawcze, tak wirusologia wykorzystuje metody biologii molekularnej do rozwiązywania swoich problemów. Technologia komputerowa zajmuje się analizą informacji genetycznej, w związku z czym pojawiły się nowe obszary genetyki molekularnej, które czasami uznawane są za dyscypliny szczególne: bioinformatykę, genomikę i proteomikę.
Historia rozwoju
Do tego przełomowego odkrycia przyczyniła się długa faza badań nad genetyką i biochemią wirusów i bakterii.
W 1928 roku Frederick Griffith po raz pierwszy wykazał, że ekstrakt z bakterii chorobotwórczych zabitych ciepłem może przenieść cechę chorobotwórczości na bakterie łagodne. Badania transformacji bakterii doprowadziły dalej do oczyszczenia czynnika chorobotwórczego, który wbrew oczekiwaniom okazał się nie białkiem, a kwasem nukleinowym. Sam kwas nukleinowy nie jest niebezpieczny, niesie jedynie geny decydujące o patogeniczności i innych właściwościach mikroorganizmu.
W latach 50. XX wieku wykazano, że bakterie mają prymitywny proces płciowy, potrafią wymieniać pozachromosomalne DNA, plazmidy. Odkrycie plazmidów, a także transformacje stworzyły podstawę technologii plazmidowej powszechnej w biologii molekularnej. Kolejnym ważnym odkryciem dla metodologii było odkrycie na początku XX wieku wirusów bakteryjnych, bakteriofagów. Fagi mogą również przenosić materiał genetyczny z jednej komórki bakteryjnej do drugiej. Zakażenie bakterii przez fagi prowadzi do zmiany składu bakteryjnego RNA. Jeśli bez fagów skład RNA jest podobny do składu DNA bakteryjnego, to po infekcji RNA staje się bardziej podobne do DNA bakteriofaga. W ten sposób odkryto, że o strukturze RNA decyduje struktura DNA. Z kolei tempo syntezy białek w komórkach zależy od ilości kompleksów RNA-białko. Tak to zostało sformułowane centralny dogmat biologii molekularnej: DNA ↔ RNA → białko.
Dalszemu rozwojowi biologii molekularnej towarzyszył zarówno rozwój jej metodologii, w szczególności wynalezienie metody określania sekwencji nukleotydów DNA (W. Gilbert i F. Sanger, Nagroda Nobla w dziedzinie chemii w 1980 r.), jak i nowe odkrycia w dziedzinie badań nad strukturą i funkcjonowaniem genów (patrz. Historia genetyki). Już na początku XXI wieku uzyskano dane dotyczące pierwotnej struktury całego ludzkiego DNA oraz szeregu innych, najważniejszych dla medycyny organizmów, m.in. Rolnictwo i badania naukowe, które doprowadziły do powstania kilku nowych dziedzin biologii: genomiki, bioinformatyki itp.
Zobacz też
- Biologia molekularna (czasopismo)
- Transkryptomika
- Paleontologia molekularna
- EMBO – Europejska Organizacja Biologii Molekularnej
Literatura
- Piosenkarz M., Berg P. Geny i genomy. - Moskwa, 1998.
- Stent G., Kalidar R. Genetyka molekularna. - Moskwa, 1981.
- Sambrook J., Fritsch E.F., Maniatis T. Klonowanie molekularne. - 1989.
- Patruszew L.I. Ekspresja genów. - M.: Nauka, 2000. - 000 s., il. ISBN 5-02-001890-2
Spinki do mankietów
Fundacja Wikimedia. 2010 .
- Rejon Ardatowski w obwodzie Niżnym Nowogrodzie
- Rejon Arzamas w obwodzie Niżnym Nowogrodzie
Zobacz, co „biologia molekularna” znajduje się w innych słownikach:
BIOLOGIA MOLEKULARNA- uczy się podstaw. właściwości i przejawy życia na poziomie molekularnym. Najważniejsze kierunki w M.b. to badania strukturalnej i funkcjonalnej organizacji aparatu genetycznego komórek oraz mechanizmu wdrażania informacji dziedzicznej ... ... Biologiczny słownik encyklopedyczny
BIOLOGIA MOLEKULARNA- bada podstawowe właściwości i przejawy życia na poziomie molekularnym. Dowiaduje się, w jaki sposób i w jakim stopniu wzrost i rozwój organizmów, przechowywanie i przekazywanie informacji dziedzicznych, przemiana energii w żywych komórkach i inne zjawiska wynikają z... Wielki słownik encyklopedyczny
BIOLOGIA MOLEKULARNA Nowoczesna encyklopedia
BIOLOGIA MOLEKULARNA- BIOLOGIA MOLEKULARNA, biologiczne badanie struktury i funkcji CZĄSTECZEK tworzących organizmy żywe. Główne kierunki studiów to fizyka i Właściwości chemiczne białka i KWASY NUKLEINOWE, takie jak DNA. Zobacz też… … Naukowy i techniczny słownik encyklopedyczny
Biologia molekularna- część biol., która bada podstawowe właściwości i przejawy życia na poziomie molekularnym. Dowiaduje się, jak i w jakim stopniu wzrost i rozwój organizmów, przechowywanie i przekazywanie informacji dziedzicznych, przemiana energii w żywych komórkach oraz... ... Słownik mikrobiologii
Biologia molekularna- — Tematy biotechnologii PL biologia molekularna … Podręcznik tłumacza technicznego
Biologia molekularna- BIOLOGIA MOLEKULARNA, bada podstawowe właściwości i przejawy życia na poziomie molekularnym. Dowiaduje się, jak i w jakim stopniu wzrost i rozwój organizmów, przechowywanie i przekazywanie informacji dziedzicznych, przemiana energii w żywych komórkach oraz... ... Ilustrowany słownik encyklopedyczny
Biologia molekularna- nauka, która za swoje zadanie stawia sobie poznanie natury zjawisk życiowych poprzez badanie obiektów i układów biologicznych na poziomie zbliżonym do poziomu molekularnego, a w niektórych przypadkach osiągającego tę granicę. Ostatecznym celem tego jest…… Wielka encyklopedia radziecka
BIOLOGIA MOLEKULARNA- bada zjawiska życia na poziomie makrocząsteczek (rozdz. białka arr i kwasy nukleinowe) w strukturach bezkomórkowych (rybosomy itp.), w wirusach, a także w komórkach. cel M. ustalenie roli i mechanizmu działania tych makrocząsteczek na podstawie ... ... Encyklopedia chemiczna
Biologia molekularna- bada podstawowe właściwości i przejawy życia na poziomie molekularnym. Dowiaduje się, jak i w jakim stopniu wzrost i rozwój organizmów, przechowywanie i przekazywanie informacji dziedzicznych, przemiana energii w żywych komórkach i inne zjawiska... ... słownik encyklopedyczny
Książki
- Biologia molekularna komórki. Książka problemów, J. Wilson, T. Hunt. Książka autorów amerykańskich stanowi dodatek do drugiego wydania podręcznika „Biologia molekularna komórki” autorstwa B. Albertsa, D. Braya, J. Lewisa i in. Zawiera pytania i zadania, których celem jest pogłębienie. ..
Biologia molekularna, nauka, która stawia sobie za zadanie poznanie natury zjawisk życiowych poprzez badanie obiektów i układów biologicznych na poziomie zbliżonym do poziomu molekularnego, a w niektórych przypadkach sięgającego tej granicy. Ostatecznym celem w tym przypadku jest wyjaśnienie, w jaki sposób i w jakim stopniu charakterystyczne przejawy życia, takie jak dziedziczność, rozmnażanie się własnego rodzaju, biosynteza białek, pobudliwość, wzrost i rozwój, przechowywanie i przekazywanie informacji, przemiany energetyczne, mobilność, itp. wynikają ze struktury, właściwości i interakcji cząsteczek substancji biologicznie ważnych, przede wszystkim dwóch głównych klas biopolimerów wielkocząsteczkowych – białek i kwasów nukleinowych. Charakterystyczną cechą M.b. - badanie zjawisk życia na obiektach nieożywionych lub tych, które charakteryzują się najbardziej prymitywnymi przejawami życia. Są to twory biologiczne z poziomu komórkowego i poniżej: organelle subkomórkowe, takie jak izolowane jądra komórkowe, mitochondria, rybosomy, chromosomy, błony komórkowe; dalej - systemy stojące na granicy przyrody ożywionej i nieożywionej - wirusy, w tym bakteriofagi, a kończąc na cząsteczkach krytyczne komponenty materia żywa - kwasy nukleinowe i białka.
Podstawę, na której rozwinął się M., położyły takie nauki jak genetyka, biochemia, fizjologia procesów elementarnych itp. Zgodnie z początkami swojego rozwoju M. b. jest nierozerwalnie związana z genetyką molekularną, która nadal stanowi ważną część
Charakterystyczną cechą M.b. jest jego trójwymiarowość. Istota M.b. M. Perutz widzi to w interpretacji funkcji biologicznych w kategoriach struktury molekularnej. M. ur. stawia sobie za zadanie uzyskanie odpowiedzi na pytanie „jak”, znając istotę roli i udziału całej struktury cząsteczki, oraz na pytania „dlaczego” i „dlaczego”, dowiedziawszy się z jednej strony , związek między właściwościami cząsteczki (ponownie, przede wszystkim białek i kwasów nukleinowych) a funkcjami, które pełni, a z drugiej strony rolą takich indywidualnych funkcji w ogólnym kompleksie przejawów aktywności życiowej.
Najważniejsze osiągnięcia biologii molekularnej. Oto niepełna lista tych osiągnięć: ujawnienie struktury i mechanizmu biologicznej funkcji DNA, wszystkich typów RNA i rybosomów, ujawnienie kodu genetycznego; odkrycie odwrotnej transkrypcji, czyli syntezy DNA na matrycy RNA; badanie mechanizmów działania barwników oddechowych; odkrycie struktury trójwymiarowej i jej funkcjonalnej roli w działaniu enzymów, zasady syntezy matrycy i mechanizmów biosyntezy białek; ujawnienie budowy wirusów i mechanizmów ich replikacji, pierwotnej i częściowo przestrzennej struktury przeciwciał; izolacja poszczególnych genów, chemiczna, a następnie biologiczna (enzymatyczna) synteza genów, w tym ludzkich, poza komórką (in vitro); przenoszenie genów z jednego organizmu do drugiego, w tym do komórek ludzkich; szybko postępujące rozszyfrowanie budowy chemicznej coraz większej liczby pojedynczych białek, głównie enzymów, a także kwasów nukleinowych; odkrycie zjawisk „samoorganizacji” niektórych obiektów biologicznych o coraz większej złożoności, począwszy od cząsteczek kwasów nukleinowych, a skończywszy na wieloskładnikowych enzymach, wirusach, rybosomach itp.; wyjaśnienie allosterycznych i innych podstawowych zasad regulacji funkcji i procesów biologicznych.
Problemy biologii molekularnej. Wraz z określonymi ważnymi zadaniami M. miałby. (znajomość praw „rozpoznania”, samoorganizacji i integracji) aktualnym kierunkiem poszukiwań naukowych na najbliższą przyszłość jest rozwój metod pozwalających na rozszyfrowanie struktury, a następnie trójwymiarowej, przestrzennej organizacji wielkocząsteczkowych kwasy nukleinowe. Wszystkie najważniejsze metody, których zastosowanie zapewniło pojawienie się i sukces M. b., zostały zaproponowane i opracowane przez fizyków (ultrawirowanie, analiza dyfrakcji promieni rentgenowskich, mikroskopia elektronowa, jądrowy rezonans magnetyczny itp.). Prawie wszystkie nowe fizyczne podejścia eksperymentalne (na przykład zastosowanie komputerów, synchrotronu lub bremsstrahlunga, promieniowania, technologii laserowej i innych) otwierają nowe możliwości dogłębnego badania problemów M. b. Do najważniejszych zadań o charakterze praktycznym, na które odpowiedzi oczekuje się od M. b., należy przede wszystkim problem molekularnych podstaw wzrostu złośliwego, a następnie – sposoby zapobiegania, a być może i przezwyciężania chorób dziedzicznych – ” choroby molekularne”. Duże znaczenie będzie miało wyjaśnienie molekularnych podstaw katalizy biologicznej, czyli działania enzymów. Wśród najważniejszych nowoczesne trendy M. ur. powinna obejmować chęć rozszyfrowania molekularnych mechanizmów działania hormonów, substancji toksycznych i leczniczych, a także poznania szczegółów budowy molekularnej i funkcjonowania takich struktur komórkowych jak błony biologiczne biorące udział w regulacji procesów penetracji i transport substancji. Bardziej odległe cele M. b. - znajomość natury procesów nerwowych, mechanizmów pamięci itp. Jedna z ważniejszych powstających sekcji M.b. - tak zwana. inżynieria genetyczna, która za swoje zadanie stawia celowe działanie aparatu genetycznego (genomu) organizmów żywych, począwszy od drobnoustrojów i niższych (jednokomórkowych), a skończywszy na człowieku (w tym drugim przypadku przede wszystkim w celu radykalnego leczenia choroby dziedziczne i korekcja wad genetycznych).
Najważniejsze kierunki MB:
- Genetyka molekularna - badanie strukturalnej i funkcjonalnej organizacji aparatu genetycznego komórki oraz mechanizmu wdrażania informacji dziedzicznej
– Wirusologia molekularna – nauka o molekularnych mechanizmach interakcji wirusów z komórkami
– Immunologia molekularna – badanie wzorców reakcji immunologicznych organizmu
– Molekularna biologia rozwoju – nauka o pojawianiu się różnorodności komórek w trakcie indywidualnego rozwoju organizmów i specjalizacji komórek
Główne obiekty badań: Wirusy (w tym bakteriofagi), Komórki i struktury subkomórkowe, Makrocząsteczki, Organizmy wielokomórkowe.